
长期以来,研究人员一直试图了解和预测动物对压力刺激的反应。自引入稳态概念以来,已经提出了各种模型框架来描述动物保持这种稳定生理状态所必需的条件以及离开这种状态的后果。Romero等人(Horm Behav 55(3): 375-389, 2009)引入了反应性范围模型,为应激反应提供了一个新的概念框架,该模型假设动物对应激刺激的耐受能力可能随着时间的推移而降低。我们用常微分方程组给出了反应作用域模型的数学表达式,并证明了该模型能够再现现有的实验数据。我们还提供了一种实验方法,可用于验证模型以及对模型的几个潜在补充。如果未来的实验提供必要的数据来估计模型的参数,这里提出的模型可用于对应激反应期间的生理介质水平进行定量预测,并预测稳态过载的发生。
根据Levine(2005)的说法,关于内稳态及其破坏的概念,也就是现在所说的压力,至少可以追溯到早期的希腊医生,而我们对压力的现代观点很大程度上可以追溯到Cannon(1932)和Selye(1946)。为动物的压力提供一个定义明确且被广泛接受的定义的尝试已经进行了很长时间,如果刺激引起压力反应,并且压力反应作为对压力源的生理或行为反应,那么将刺激定义为压力源的周期性本质就会产生许多模糊性(Levine 2005;Romero et al. 2009)。最近,Sterling和Eyer(1988)提出了异稳态模型,试图从能量获取和使用的角度重新定义压力的概念。随后,McEwen和Wingfield(2003)利用适应平衡的概念,通过觅食成功、恶劣天气和栖息地变化等生态因素来解释糖皮质激素等生理介质水平。
Romero等人(2009)引入了反应性作用域模型,以进一步完善异稳态模型,同时解决其弱点。与着重于资源分配和支出的非稳态模型不同,反应性范围模型允许对压力或压力事件的概念进行广泛的解释,从而考虑到动态平衡的变化,甚至对动态平衡变化的预期。广义地说,反应作用域模型引入了两个考虑应力的新成分。首先,压力源或压力事件的概念,即导致压力的刺激,被允许包括对环境压力源的预期反应。第二是在传统的内稳态模型中加入了同名的反应性内稳态范围。反应性内稳态范围被认为是一个动态缓冲带,代表了传统内稳态和内稳态过载之间的暂时弹性,当动物经历应激事件导致内稳态外的中介水平时,该缓冲带会暂时或永久地收缩。作者使用“磨损”一词来指由于维持应激反应的累积成本而导致的反应性稳态范围的缩小。作者还指出,在这种情况下,“磨损”一词的使用不应该与损伤和老化的缓慢积累联系在一起。
自反应性范围模型引入以来,许多研究都根据该模型来解释结果(Aguilar et al. 2016;Beattie et al. 2023a;Charpentier et al. 2018;Crespi et al. 2013;Eguizábal等人,2022;Gabriel et al. 2018;Houtz et al. 2022;Howell and Sanchez 2011;Leishman et al. 2022;Lima et al. 2022;Pahuja和Narayan 2021;Roast等人,2019;罗梅罗2012;Schoenle等人,2018年)。另一方面,相对较少的研究,包括Gormally等人(2019b)、Beattie等人(2023b)、Gormally等人(2019a)、DuRant等人(2016b)和DuRant等人(2016a),都以验证反应性范围模型为具体目标,并取得了不同程度的成功。一般来说,这些研究在野外捕获的鸟类群体中引起应激反应,允许各组恢复不同的时间长度,然后诱导新的应激反应,假设恢复时间较长的组将有足够的时间从反应性稳态中恢复过来,从而使应激反应比恢复时间较短的组减少。然而,由于当前版本的反应范围模型纯粹是概念性的,这些研究人员没有办法确定应力事件或恢复的必要持续时间,以确保假设的磨损和撕裂是可观察到的。
正如Grindstaff et al.(2022)、Servedio et al.(2014)和Zavala et al.(2019)所指出的,数学模型可以成为研究理论生物学和内分泌学理论的有力工具。在Luttbeg et al.(2021)、Taborsky et al.(2021)和Taborsky et al.(2022)中,作者使用数学模型来探索应激反应的演变以及修复率如何影响基础水平。数学框架和模型已经被用于垂体-肾上腺(HPA)轴、应激和应激反应的其他研究。自Dempsher等人(1984)提供的开创性工作以来,使用数学模型来研究下丘脑轴本身已经引起了极大的兴趣。Lightman(2008)、Goel等人(2014)、Nicolaides等人(2017)、Tsigos和Chrousos(2002)、Lightman和Conway-Campbell(2010)、Vinther等人(2011)、Gudmand-Hoeyer等人(2014)、Spiga等人(2015)、Zavala等人(2019)和Hosseinichimeh等人(2015)都对这一主题进行了综述。Hosseinichimeh等人(2015)特别提供了14个已发表模型的比较,并试图将它们与具有不同结果的数据相匹配,然后继续提供最有希望的模型的更新形式。自这些评论以来,其他人,如Rao和Androulakis(2019)和stanojeviki等人(2018),继续努力探索这些模型并讨论进展。到目前为止,构建的模型试图捕捉HPA轴的复杂本质,该轴由分子相互作用系统、反应-运输过程和途径组成,使用常微分方程或延迟微分方程系统来描述基础和应激诱导的人类皮质醇水平或大鼠皮质酮水平。然而,正如stanojeviki等人(2018)指出的那样,虽然HPA轴的数学建模领域丰富且不断发展,但“对于核心反馈机制仍然没有共识或共同的表示。”此外,这些模型通常是用特定的数据集构建和校准的,模型的预测没有在其他已发表的数据集上进行测试。此外,虽然对这些模型的分析包括了对所描述系统的急性压力事件进行的硅实验,但它们并没有探索系统应对长期或严重压力的能力的退化。这些模型也可能在其设计的物种之外几乎没有应用,并且参与应激反应过程的分子可能因物种而异(Taborsky et al. 2021)。最后,虽然HPA轴的现有模型是复杂系统的简化,但对于那些背景不包括大量数学的研究人员来说,它们可能是不可接近的(Fawcett和Higginson 2012)。
在当前的表述中,反应作用域模型的一个弱点是它是一个图形化的概念框架。也就是说,到目前为止,还没有提供反应作用域模型的定量表示。因此,无法预测实验结果,从而无法客观地验证模型。Grindstaff et al.(2022)指出,数学模型的制定本身并不能验证反应作用域模型。确认概念模型将需要经验主义和机械论模型的联合工作,就像这里展示的那样。通过描述反应范围模型的数学模型,可以利用回归技术和实验数据对数学模型的参数进行估计,然后该模型可以用于预测特定情况下的生理介质水平,这种预测可以反过来与实验数据进行比较。如果发现模型和数据很一致,我们可以假设数学模型准确地捕捉了应力反应过程。如果没有,则可以根据需要调整模型。
我们为反应作用域模型提供了一个定量框架,以及它的几个组件。在2.1节中,我们开始描述一般的应力事件,并将这些应力事件用数学方法表示为时间的函数。在第2.2节中,我们提供了受昼夜节律和季节变化影响的中介水平的时间依赖形式,然后将反应范围模型描述为一个常微分方程系统,该系统描述了反应范围模型中范围之间的边界。接下来,我们将提供几个例子来说明这个数学模型的灵活性。在第3节中,我们使用数学模型来复制实证研究的结果,并对数学模型进行了广泛的分析。在第4节中,我们提供了一种经验方法来确定模型的准确性。最后,在第5.1节中,我们提供了对反应作用域模型的几个可能的补充,并讨论了它的弱点。
Fischer和Romero(2013)、McEwen(2003)、McEwen和Wingfield(2003)以及Romero等人(2009)都试图描述应激对生理介质的影响,如糖皮质激素、抗体滴度、儿茶酚胺和心率等。在反应性范围模型中,Romero等人(2009)假设McEwen(2003)和McEwen和Wingfield(2003)提出的生理介质,如行为改变、免疫功能介质和HPA轴介质,可根据范围分为四类:预测性内稳态、反应性内稳态、内稳态过载和内稳态失效。我们在这里采用相同的术语。当动物能够补偿任何由季节和昼夜变化引起的正常波动以及可能与适应平衡相关的正常觅食行为时,中介值处于可预测的内稳态范围内。Romero et al.(2009)引入了反应性稳态范围。反应性内稳态范围内的中介值超出了预测内稳态范围所代表的正常波动,但仍然足够低,暂时避免进入内稳态过载。预测性内稳态和反应性内稳态这两个术语来自Moore-Ede(1986),它们共同代表了动物的正常反应范围。超过反应性内稳态范围的中介物水平被认为处于内稳态过载范围,并将诱发病理状态。Romero等人(2009)将最后一个范围描述为内稳态失效,低于预测的内稳态范围,表示动物无法维持或调节正常的介质水平,随后迅速死亡。图1给出了无应力事件时反应作用域模型的图形表示,其中预测稳态范围、反应稳态范围和稳态过载范围之间的阈值分别标记为P(t)和R(t)。有关这些值的进一步讨论,请参见第2.2节。
图1
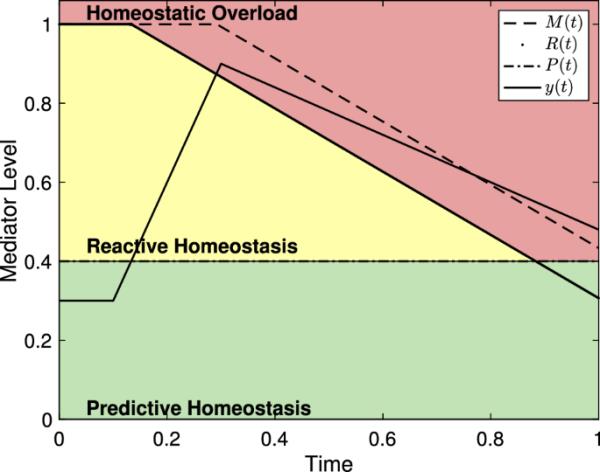
反应性范围模型中的中介水平和(8)中没有压力事件的系统的一般例子,用于重复Romero等人(2009)给出的描述反应性范围的数字。请注意,反应性内稳态范围的大小(R(t)和P(t)之间的距离)正在缩小,这是由于预测内稳态范围大小的长期季节性变化。中介值的波动是为了模拟昼夜变化
反应性范围模型的一个重要方面是损耗:与每个分类相关联的范围的大小可以在响应压力事件时临时或永久地改变。“大小”是指中介y轴上的范围之间的阈值之间的距离。这种磨损的结果是,在诱导体内平衡超载之前,动物可以耐受一段时间内升高的介质水平,模仿弹性,并且相同的应激反应可能在未来更快地导致体内平衡超载。Romero et al.(2009)认为,“进入稳态过载有两种方式:介质的浓度或水平超出正常反应范围;或者介质的浓度或水平在较长时间内保持在反应性稳态范围内。”
我们在下面介绍我们的模型,首先讨论并给出应力事件和时间表的数学表示,然后是反应范围模型的三个主要特征范围之间的阈值。
Romero et al.(2009)将应激事件定义为动物环境中不可预测的事件,应激反应是对该事件的生理反应。他们强调压力反应的确切行为取决于所讨论的中介或行为。此外,应激事件或反应可能有多种原因,包括McEwen和Wingfield(2003)所描述的适应负荷、捕食,甚至可能是对应激源事件的预期。在这里,我们忽略了刺激事件和反应之间的区别,并使用术语应激事件来指代生理介质水平的任何变化,而不管原因是环境刺激、适应负荷还是心理原因。为了描述压力事件或一系列压力事件中的中介水平,我们使用如下所述的压力表
(1)
其中n是所考虑的压力事件的数量,是压力事件的压力响应的大小,是捕获压力事件期望行为的函数(例如上下调节时间,增加和减少速率等),和是指示事件时间和持续时间的参数。应该注意的是,不同的中介在任何给定的情况下都会有不同的反应,压力事件和随后的压力计划旨在捕捉单个中介随时间的行为。因此,如果考虑几个介质,它们各自的应力表应该绘制在不同的轴上。
我们在下面提供了三种可能的形式,它们的图形如图2所示。首先,Romero等人(2009)提供的图形模型描述了使用
(2)
其中和是严格的正常数。在这里,表示压力事件的开始,并表示事件的持续时间。这种形式对演示目的是有用的,但可能提供了压力反应的过于简化的表示,因为它将导致介质水平瞬间上下调节。
Rich和Romero(2005)记录了受诱导应激的鸟类的皮质酮水平。这些数据表明,应激反应始于应激事件的开始,并在整个应激事件中被上调。在应激事件结束时,中介体在一段比应激事件更长的时间内被下调。用给出的表达式更好地表示这个过程
(3)
在哪里和。的值应该与压力事件的开始一致(或者如果刺激和反应之间的延迟时间已知,则可以调整),并表示压力事件或上调期的持续时间。该参数表示中介器下调所需的时间。例如Rich和Romero(2005)研究表明,CORT的下调时间是上调时间的3 - 4倍。
其他选项可能更适合于视情况而定。例如,
(4)
其中表示压力事件的中间时间将提供压力事件的连续可微分版本,可以更准确地捕捉上下调节的性质。然而,在Eq.(4)中并不能直接表示压力事件或反应的持续时间。也就是说,较大的值意味着较长的持续时间,但事件和响应的持续时间都不等于。图2比较了这里提供的压力事件的三种描述。
图2
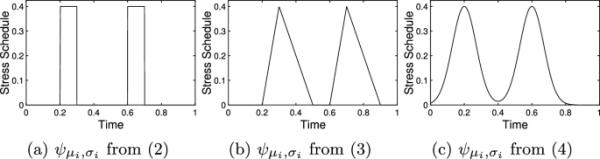
(a)应力表的形式见(2)。应力表中,、、和使用不同的形式表示。这些时间表是独立于考虑生理介质而提出的,因此独立轴上的值在这里没有意义。b。请注意,每个事件都从适当的值开始。的值表示事件发生的中间时间
反应性作用域模型的一个定义特征是(Romero et al. 2009)所说的“损耗”。与使用“磨损”来指长期、缓慢的损伤和老化的积累不同(McEwen 1998),磨损是指反复或长时间的应激反应可能导致动物在应激反应期间反应性稳态范围的减小,从而使未来相同强度的应激事件有可能导致稳态过载。我们通过改变反应作用域模型的特征范围之间的阈值曲线来模拟这种行为。
由于反应范围模型对生理中介水平的反应发生变化,我们首先描述在时间t到y(t)时所考虑的中介水平,其中
(5)
和分别是季节和昼夜变化的振幅,分别是季节和昼夜变化的周期,是生理中介的基础水平(必须选择足够大以确保y(t)是非负的),s(t)是(1)中定义的应激时间表。因此,中介水平y(t)是生理因素和应激时间表共同驱动的。
接下来,我们描述了预测内稳态范围和反应性内稳态范围之间的初始阈值曲线P(t)
(6)
其中,、、与上文相同,表示由于可预测的活动而导致的中介级别的变化。
然后,我们用R(t)表示反应性稳态和稳态过载之间的边界,并通过允许R(t)响应y(t)而变化来描述反应性范围的假设磨损。由于反应作用域模型假设,如果应力事件未能引起稳态过载,反应作用域范围将恢复到刺激前的大小,因此我们引入一个新的量M(t)来表示应力事件结束时R(t)可以恢复到的最大值。根据Romero et al.(2009)用我们的符号表示,对于所有的t, R(t)应该暂时减小,如果应该永久减小,当增加到M(t)。进一步,M(t)在。我们用Heaviside函数表示
(7)
然后,
(8)
在那里;、、和都是严格正的;和。(8)给出的系统模拟了Romero et al.(2009)给出的图,如图1所示。在没有应力事件的情况下,对于所有的t值和它们的曲线重叠。这里,,,,。注意,内稳态失效范围将出现在预测的内稳态范围之下,但为了简单起见,在后面的图中省略了。为了这项工作,除非另有说明,否则我们假定。在第5.1节中,我们提供了对基本模型的几种可能的补充,并将重点放在基本模型的特征上。
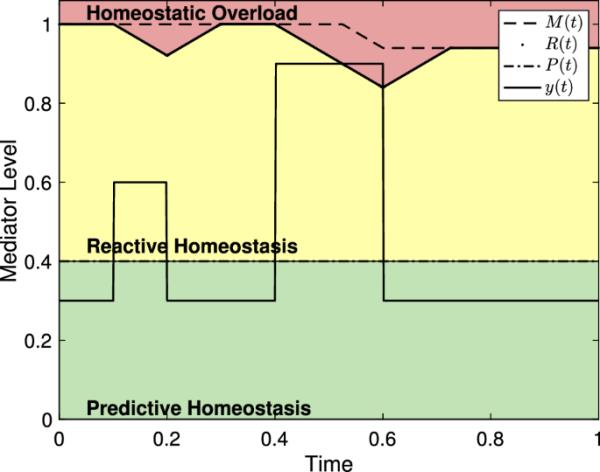
为了证明模型的特征,我们考虑了一些独立于任何特定实验或中介水平的例子。我们设置了从0到1的独立轴,其中1可以被认为是最大效果的水平(类似于药理学(Derendorf et al. 2020))。同样,相关轴被缩放为1以表示实验持续时间,除非另有说明,否则我们继续这种惯例。图3显示了反应作用域模型的几种表示形式,以及各种选项。对于每个选择,其时间长度不同,导致R(t)中的行为不同。因为对于每个图中的所有t,对于每个图。
对于图3和模型的所有其余描述,将自稳态失效范围删除,以关注图的相关部分。此外,除第3节中的数字外,本研究中的数字旨在描述任意生理介质与稳态过载(红色)和反应性稳态(黄色)范围之间的定性关系。因此,轴的缩放实际上是任意的和不相关的。
图3
反应作用域模型中的中介水平和稳态范围。由于没有特定的中介呈现,依赖轴上的值1对应于最大效果的中介水平。同样,独立轴上的值可以看作是实验持续时间的比例。A - c压力表,其中,,和。这些范围的颜色与图1中的范围一致。选B。请注意,每个事件都从适当的值开始。的值表示事件发生的中间时间
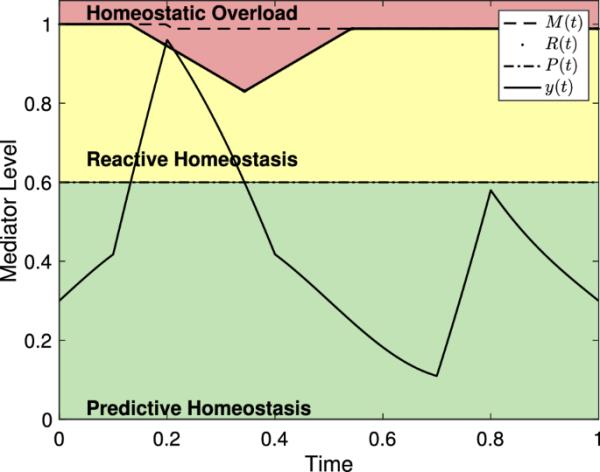
图4显示了反应性范围以及由具有两个压力事件的压力计划驱动的中介。由于第一次应力事件不会引起稳态过载,因此R(t)的值会暂时降低,而M(t)不会。因此,R(t)恢复到原来的最大值。第二次应力事件持续足够长的时间,使反应范围减小,导致稳态过载,而介质水平没有进一步增加。因此,M(t)的值永久减小。因此,阈值R(t)永远无法恢复到原始值。
图4
具有中介级别的反应性范围模型,由具有两个压力事件的压力计划驱动。这里是,,,,,,和。第一次应力事件不引起稳态过载,因此M(t)的值保持固定,而R(t)减小后恢复到M(t)的值。在第二次应力事件中,y(t)的升高导致R(t)降至y(t)以下,导致稳态过载,导致M(t)永久下降。
图5显示了反应范围以及由压力计划和经历昼夜变化驱动的介质。在这两种情况下,压力事件的大小是相同的,但第一种情况会导致体内平衡超载,因为它发生在基础水平达到峰值时。因此M(t)永久减小。尽管具有相同的量级,但第二次应激事件不会导致中介水平增长到预测稳态范围以上,因此R(t)不会减小。
图5
反应范围与中介驱动的压力时间表和昼夜变化。这里,,,和。对于压力时间表,,,,和。取自(3)with
摘要
1 介绍
2 该模型
3.实验结果的重复性
4 模型验证
5 讨论
6 总结
参考文献
致谢
作者信息
道德声明
搜索
导航
#####
许多实证研究使用反应性范围模型解释了结果(Aguilar et al. 2016;Beattie et al. 2023a;Charpentier et al. 2018;Crespi et al. 2013;Eguizábal等人,2022;Gabriel et al. 2018;Houtz et al. 2022;Howell and Sanchez 2011;Leishman et al. 2022;Lima et al. 2022;Pahuja和Narayan 2021;Roast等人,2019;罗梅罗2012;Schoenle et al. 2018)。然而,只有Beattie et al. (2023b)、Gormally et al. (2019b)、Gormally et al. (2019a)、DuRant et al. (2016b)和DuRant et al. (2016a)等少数研究明确目的是研究反应性范围模型。由于这些研究包括使用已知的可接受的情况来引发应激反应,定期测量与应激反应相关的生理介质,并为使用该模型的结果的再创造提供充足的信息,因此我们在这里重点关注后一组。这些研究主要集中在鸟类的皮质酮水平上。皮质酮(CORT)是主要的鸟类糖皮质激素(Holmes and Phillips 1976),儿茶酚胺也被发现是应激反应的有效指标,很适合反应性范围模型(Fischer and Romero 2013)。我们复制了DuRant等人(2016a)所显示的与CORT相关的一些结果,其中CORT水平在25天内测量了四次。由于研究中的测量频率太低,无法捕捉昼夜变化,因此我们将以Breuner等人(1999)发现的基础水平为基础,该研究发现甘贝尔白冠麻雀(斑鸠白冠麻雀)的基础皮质醇水平每天都在变化,活动期前的最大基础值低于30 ng/ml。我们将假设类似的模式适用于DuRant等人(2016a)使用的家麻雀(Passer domesticus)。由于实验时间相对较短,我们将忽略季节变化。
为了模拟上面提到的情况,我们设置了ng/ml,(因此),ng/ml和。杜兰特等人(2016a)提供的数据表明,从实验的第1天到第12天,CORT水平升高,然后下降,直到第25天,也就是最后一次采血的日子。因此,我们将以(3)的形式使用单个重音事件with,以ng/ml, day, days表示。CORT水平在最后一次血液采样时还没有恢复正常,所以我们假设现在的模式继续下去。
DuRant等人(2016a)指出,人工升高CORT水平的鸟类伤口愈合能力下降,这表明这一组可能由于实验而经历了稳态过载(尽管作者指出结果没有统计学意义)。由于无法估计,,,,和给出的信息,我们通过检查确定这些值为,ng/ml。带有这些变量的模型输出如图6所示。
图6
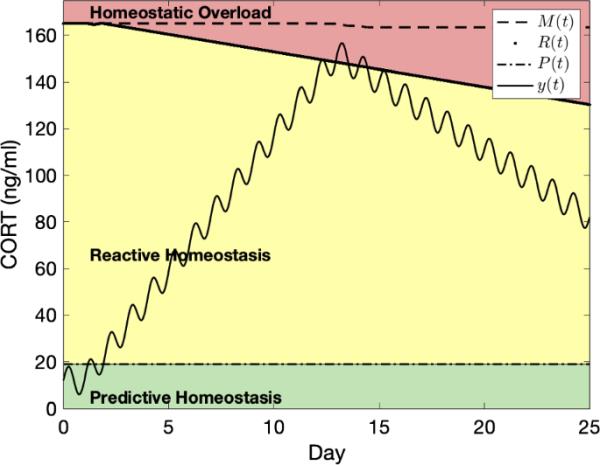
DuRant等人(2016a)对人工升高CORT水平的鸟类中介物水平的重建。振荡是由y(t)的昼夜变化引起的
我们在这里的目标不是验证或拒绝杜兰特等人的工作(2016a)。我们也不认为,使用本文提出的反应范围的数学框架来重现实证工作的能力,既验证了模型的数学公式,也验证了概念框架。相反,我们提出这个重建希望展示数学框架的灵活性和潜力。
反应性范围模型完全由中介水平y(t)驱动,而中介水平y(t)又由季节和昼夜变化以及压力计划驱动。(8)给出的系统预测R(t)和M(t)没有变化,除非。也就是说,在没有压力的情况下,反应范围没有变化。(在5.1节中,我们考虑了一个允许R(t)和M(t)随时间减小的模型版本。)在存在压力事件和中介水平升高的情况下,中介的特征在很大程度上决定了系统的行为。一个简单的、静止的钟摆可以作为这个系统的一个很好的类比。如果不动,钟摆不会发生任何变化。如果摆受到某种扰动,它就会起反应,但反应的特性取决于施加的力的具体程度。就像钟摆一样,可以通过多种方式操纵中介体水平。
对于一个轻微的压力事件,其中中介水平y(t)只是暂时超过P(t),不超过R(t),选择对模型的预测没有显著影响。(8)中的系统预测,当y(t)超过P(t)时,R(t)将减小,但这种减小与y(t)的大小和y(t)的增长速度不成比例。图7显示了使用不同震级的应力事件的反应范围系统,但对于每个时间间隔的长度是相同的。本节所有图均以无单位轴给出,可假定为表示介质实验时间的比例。因此,该模型对是否不诱导自稳态破坏的选择不敏感。
图7
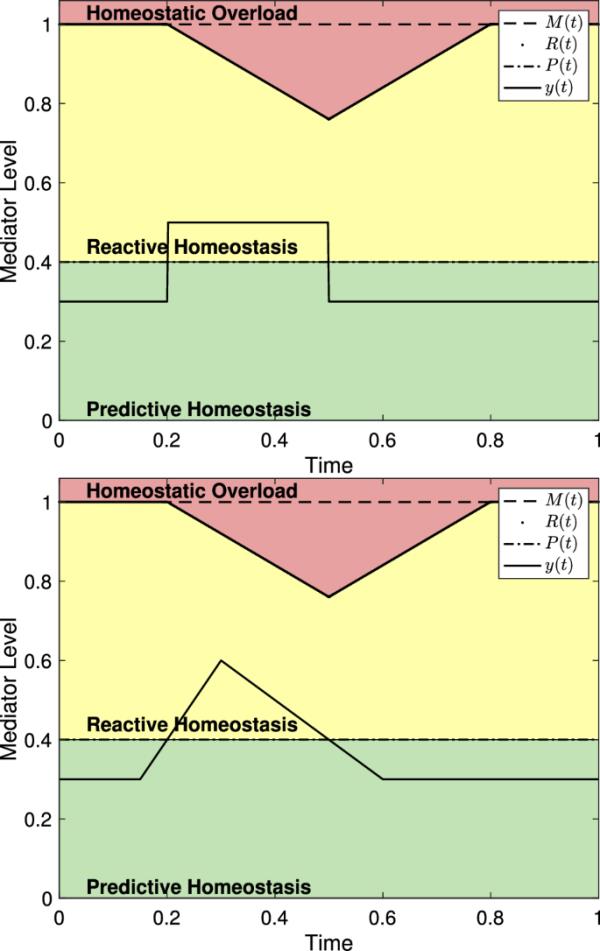
(8)中给出的两个版本的作用域系统使用了不同的函数。两者中,for和R(t)是一样的。(左)如(2)所示,其中有,,和。(右)如(3)中所示,有,,和
类似地,如果一个应力事件足够显著,使得y(t)超过R(t),那么只有导致M(t)下降的时间间隔长度。两个具有不同特征和震级的应力事件,在作用时间相同的情况下,M(t)的下降幅度相同。然而,具有不同特征的应激事件将导致中介水平在反应性稳态范围内花费不同的时间,从而导致R(t)的不同下降。图8显示了反应性作用域模型的两个版本,它们都在稳态过载中花费了相同的时间。
图8
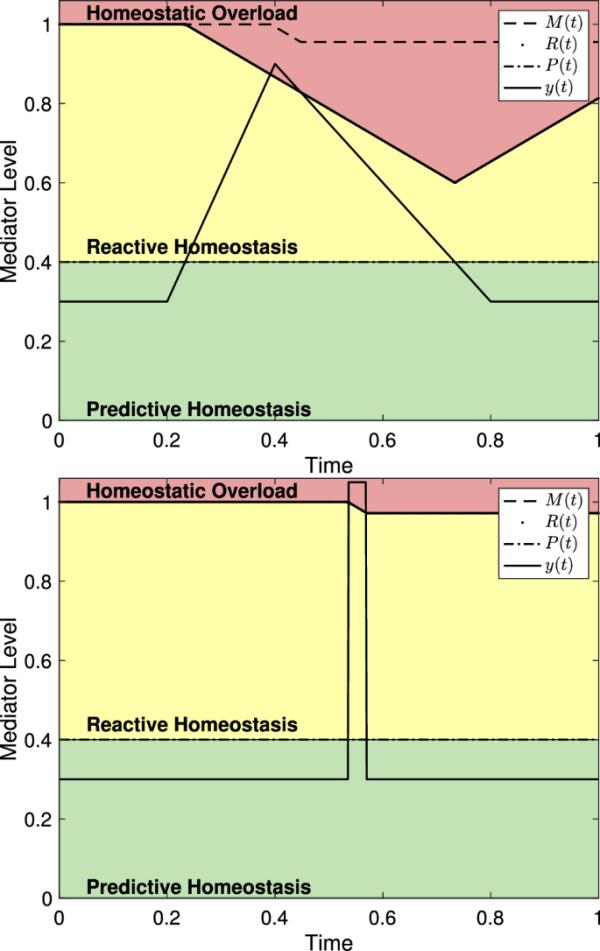
带有稳态过载的两个版本的反应作用域模型。在这两种情况下,对于大约0.033834个时间单位,因此,M(t)减少了(这里有)。(左)如(3)中带,,,和。(右)如(2)中有,,和
如图8所示,反应性范围模型非常强调压力事件的持续时间。图9显示了(8)中系统的两个版本,具有不同强度和持续时间的应力事件。由此可见,如果持续时间足够短,中介物水平的严重升高可能不会导致体内平衡超载,而中介物水平长时间升高到反应性体内平衡范围可能导致体内平衡超载。
图9
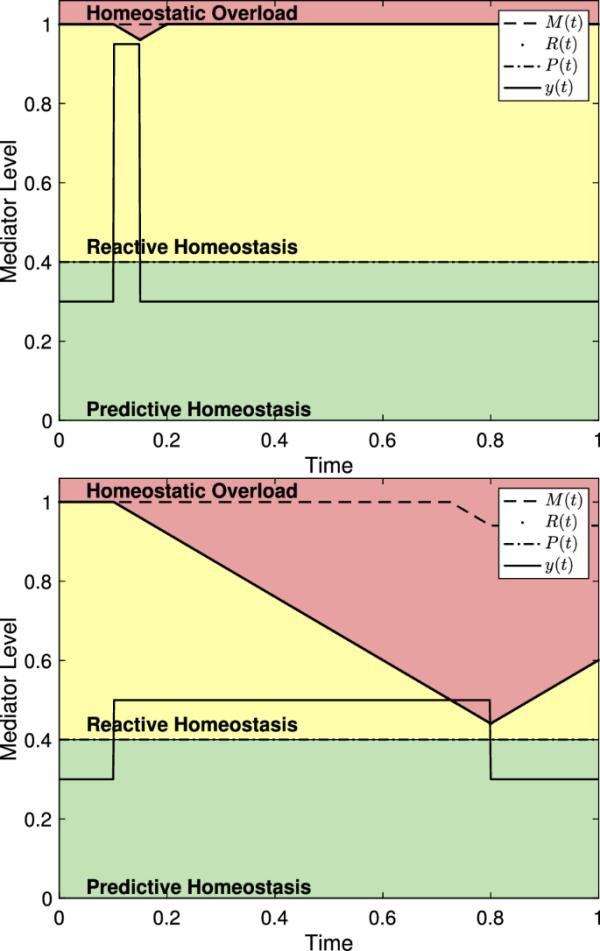
反应性范围模型显示了不同程度和持续时间的中介水平。(左)具有,且不会引起稳态过载的介质水平。(右)一个中介体水平,并导致稳态过载
生理介质的下调速率可以对R(t)的降低速率提供一些见解。如果一个动物的下调率大于,那么这个动物一旦开始就无法摆脱稳态过载。图10描述了这样一个事件。这意味着必须大于给定介质的下调率。
图10
一种生理介质,其下调发生的速率低于不能逃避稳态过载
由于反应范围模型预测应激反应期间反应性内稳态范围的下降,以及显著应激反应期间最大反应性内稳态的下降,如果重复应激事件在较短的时间间隔内引起内稳态过载,则可以验证该模型。
我们考虑一系列应力事件,每个应力事件都会引起y(t)的显著上升,足以引起稳态过载,并且间隔足够远,以允许R(t)的值恢复到M(t)的值,并假设。见图11。对于第一次应力事件,我们用时间的长度表示。在这段时间内,R(t)将下降,而y(t)继续增加,直到它们的值相交。表示曲线相交时的中介水平。因为我们知道R(t)在这段时间内减小了,我们知道。由于第一次应力事件引起稳态过载,M(t)的值将减小。若将稳态过载周期表示为,则在稳态过载周期结束时。
图11
第4节中描述的应力事件的图形描述,标注了时间范围和相应的中介水平。为了这个图的目的
同样,在第二次应力事件中,R(t)下降,直到在介质水平与y(t)相交,开始稳态过载。我们将稳态过载的持续时间表示为,并注意到M(t)在此期间下降为。因此,。这是
(9)
我们从第一次压力事件中知道,因此
(10) (11) (12)
在另一个压力事件中重复这种模式,我们可能会发现
(13)
结合(9)(13),我们发现
(14) (15) (16)
我们可以继续这个模式来找到它
(17)
这意味着n个应力事件可以用来计算总共r次。因此,如果(17)中给出的比值值在多个应激事件中是一致的,那么最大反应性稳态和反应性稳态的值必然如预测的那样线性下降。
这种方法有几个好处。首先,不需要知道的实际值。事实上,由于反应性范围模型预测在重大压力的情况下M(t)会下降,任何野生捕获的动物都可能有不同的水平。此外,任何被检查的动物个体只需要承受三次应激反应就可以进行验证,任何进一步的应激反应都可以用于进一步验证。最后,即使发现的值不一致,它们的行为也应该表明如何调整反应范围的下降。
(8)中提供的数学公式是为了尽可能模仿Romero et al.(2009)对反应作用域的描述。在本节中,我们将讨论模型的一些潜在的补充以及模型的弱点。
(8)中提出的反应性范围模型主要描述了短期情况下的应激反应。然而,动物对压力的反应可能会在其一生中发生变化(Andrews et al. 2017;Barbi et al. 2018;Haussmann and Heidinger 2015)。衰老可包括在(8)通过
(18)
其中函数A(t)描述了稳态过载的最大阈值M(t)随时间的下降。图12显示了(18)对A(t)使用不同形式的两个可能示例。作为一个例子,我们可以定义
(19)
来描述年龄和M(t)值之间的线性下降,从年龄到年龄的值。这里,如果所讨论的时间间隔是动物的整个生命周期,那么。式(19)给出的A形式的一个优点是,它允许在没有引起稳态过载的应力事件的情况下简单地确定M(t)。也就是说,对于所有的t,我们合并了期望的初始条件,那么
(20)
这允许和。这种简单性允许一旦知道A的参数就可以设置。然而,(19)中给出的A的形式不是连续的,它描述了M(t)和R(t)行为的突然变化,这在动物中可能是不现实的。
为了描述M(t)和R(t)更自然的下降,我们可以考虑M(t)在没有压力事件的情况下,如所述
(21)
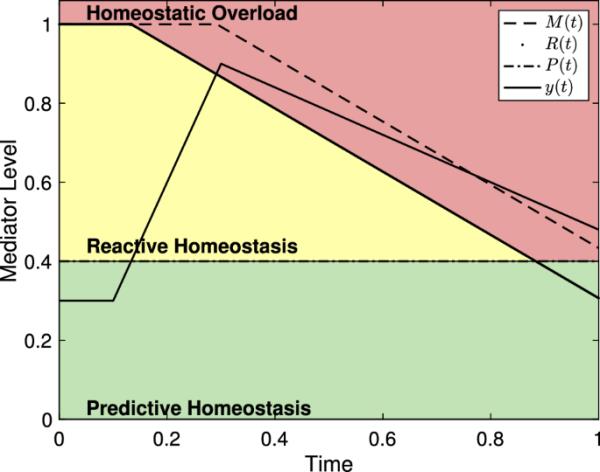
在哪儿E为的初始值米(t),表示的最终值米(t),k表示从to过渡的斜率,位置参数表示从to过渡时期的中点。例如,如果一只动物能活100天,它的生命开始于,结束于,结束于米(t)在第10天到第90天之间大致下降,那么我们会有,,和 . 一个k可以选择给出所需转换速率的值ted。然后我们可以
. 一个k可以选择给出所需转换速率的值ted。然后我们可以
(22)
图12
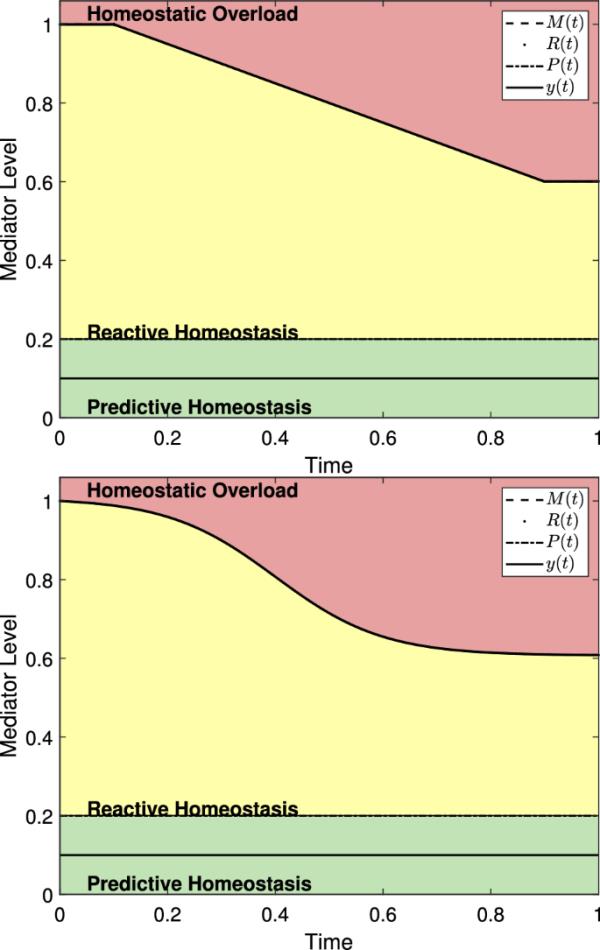
反应性范围模型显示了动物的一生归一化,包括衰老和无应激事件。(左)式(20)中描述的函数A(t)与,,,和一起使用。(右)式(22)中描述的函数A(t)与、、、和一起使用。由于没有应力,对于所有的t值和曲线重叠
图12显示了在没有压力事件的情况下,使用A(t)的两个选择,动物寿命中的M(t)。由于在反应性范围模型中包含衰老需要额外的参数,这些参数需要在动物的整个生命周期中进行观察,因此我们将进一步讨论衰老模型及其验证留到未来的工作中。
2.2节中给出的反应作用域模型版本使用Heaviside函数根据与反应作用域阈值R(t)相关的中介值y(t)来“切换”(8)中提供的系统行为。当y(t)超过R(t)时,这种转换更适合逐渐进行。给出了演示这种期望行为的函数
(23)
如(23)所示,sigmoid函数近似于(7)给出的Heaviside函数。应该注意的是,此更新仍然需要包含Heaviside函数以确保。即(8)应更新为
(24)
图13
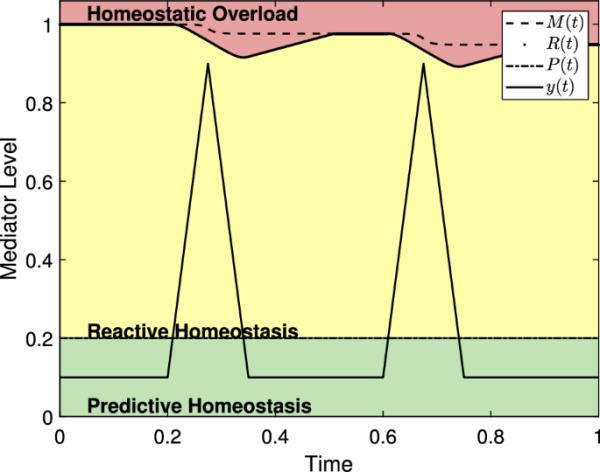
(24)中使用s型转换函数的反应作用域模型的演示。注意M(t)的值在减小,即使对于所有的t,这里,while和强调M(t)的变化
图13显示了(24)中所示的反应作用域模型。唯一显著的变化是M(t)的值减小,而这是由于强调这一行为的低选择。对于更现实的选择c(例如),(24)的行为几乎与(8)无法区分。因此,添加这个额外的函数对模型的预测几乎没有改变,而添加一个必须在实践中估计的新参数。此外,在(24)中的两个方程中,c的值可能不同。由于在模型中包含的额外复杂性,我们将不再在这里进一步考虑它,但已经提出了考虑替代方案的努力。
反应作用域模型有其局限性。在任何应激反应中都有各种生理介质在起作用。所有这些介质都在自己的稳态范围和尺度内工作,并可能导致不同水平的稳态过载(Romero et al. 2015)。因此,如果应激事件导致体内平衡超载,则可能无法确定哪个介质离开了反应性体内平衡范围并触发了体内平衡超载。越来越多的人认为,糖皮质激素可能是一种不完美的压力测量方法,测量多种介质对于理解压力反应至关重要(Creel等人,2013;Currie et al. 2010;Du et al. 2009;Jessop et al. 2013;Romero et al. 2015;Tomiyama et al. 2012)。在广泛的文献回顾之后,Dickens和Romero(2013)宣布对长期应激野生动物的内分泌特征没有共识。Gormally等人(2019b)假设,这些差异至少可以部分解释为与物种和生活史阶段相关的实验设计差异。
此外,在不同的介质、物种和个体之间,、、和甚至的值可能是不同的。因此,将实验结果应用于任何具有未知生活史的野生动物可能尤其具有挑战性。然而,如果可以建立一般范围,反应范围模型可能会为遭受污染或栖息地丧失等共同慢性压力的野生种群提供有价值的见解。
预测内稳态范围和内稳态范围之间的边界,描述为(6)中的P(t),代表了一个潜在的数学挑战,因为反应性内稳态和内稳态过载之间的边界R(t)仅在介质水平超过P(t)时下降。因此,确定第4节中描述的值需要能够根据经验确定何时发生这种情况。此外,简单地捕获野生动物并将其运送到实验室可能足以改变动物的应激生理(Dickens et al. 2009)。
Romero et al.(2009)提出的反应范围模型为动物应激反应的研究提供了一个新的框架。该框架的一个关键特征是包含了介质水平的反应性稳态范围。在反应性内稳态范围内的持续介质水平可能通过反应性内稳态范围的退化导致内稳态过载,从而允许动物生活史晚期的应激事件(或在其他近期应激事件之后)引起内稳态过载,即使在动物生活史早期(或没有其他近期应激事件)的反应水平不足以引起内稳态过载。到目前为止,反应作用域模型只是作为一个概念模型出现的。
已经进行了实验努力来测试反应范围模型的准确性(Beattie et al. 2023a;DuRant等人,2016b, a;Gormally et al. 2019b, a).这些实验试图通过对不同群体的野生鸟类造成重复的应激事件,并观察应激事件之间的恢复时间如何影响后期应激事件中的介质水平,来观察反应范围范围的退化。由于反应范围模型仅作为一个概念模型存在,实验人员无法使用该模型来确定观察反应范围退化所需的应激事件持续时间,也无法确定恢复反应范围所需的恢复时间。
我们提供了概念反应作用域模型的定量表述。已经给出了几种可能的描述符,可用于描述应力反应期间介质水平的所需曲线形状。这些描述符可以与先前收集的给定物种中特定介质的信息相结合,以描述介质水平作为包含应激事件(y(t) in(5))的时间函数。现有的数据可用于估计所需的参数,以准确描述动物的介质水平随时间的变化,因为它们每天或季节性波动。此外,我们通过提供一个常微分方程系统(8)捕捉到了反应性稳态范围的动态特性,该系统描述了中介体水平之间的阈值如何随着中介体水平的升高而变化。在估算(8)中的参数值之前,还需要做更多的工作。然而,这样的实验要么提供必要的估计,要么使反应性范围模型的当前形式无效。我们还为反应作用域模型提供了几个潜在的补充或调整,包括纳入衰老的方法。
下载原文档:https://link.springer.com/content/pdf/10.1007/s00285-023-01983-9.pdf








