
新北极有蹄类动物,如偶蹄类动物、异趾类动物和长鼻类动物,在美洲生物大交流期间抵达南美洲。其中包括骆驼科、鹿科、马科、貘科和gomphotherids。这里研究和描述了阿根廷潘潘地区更新世沉积层的新北极有蹄类动物的历史收藏。这些标本由圣地亚哥·罗斯(Santiago Roth)在19世纪收集并带到欧洲的标本组成,存放在苏黎世大学和日内瓦自然历史博物馆的古生物学收藏中。本文报道的分类群包括:platensis Notiomastodon, Lama guanicoe, Hemiauchenia paradoxa, Tayassu pecari, Morenelaphus sp., Hippidion cf. H. principale, Equus cf. E. negeus,以及其他未确定的gomphotherids, camelids, tayassuids, cerids和equids。在许多情况下,这些130多年前收集的化石的确切地层位置是不确定的。这些历史资料对该地区更新世哺乳动物区系的分类研究和古生物地理学、古生物学研究仍有重要意义。
新北极有蹄类化石,如偶蹄类、异趾类和吻肢类,在南美洲的化石记录中有很好的代表性(见Alberdi & Prado, 2002;Cassini et al., 2016;偏转,2019;Gasparini, 2013;Gasparini等,2011,2014,2017;González等人,2014;Holanda & Ferrero, 2013;母亲等人,2012,2013,2017a, b, c;Scherer, 2013,以及其中的参考文献)。它们在美洲大生物交换(GABI)期间抵达南部大陆,这一过程是由于中美洲在大约10-7 Ma建立陆桥而引发的(例如,Carrillo等人,2015;Cione et al., 2015;辛普森,1980;韦伯,1985,2006)。GABI的特征是一系列主要的迁徙“波”,主要发生在上新世-更新世,这使得北美和南美之间进行了大规模的动物交换(例如,Woodburne, 2010);尽管在中新世晚期有早期的迁移(Cione et al., 2015)。经典的解释认为GABI的开始时间为~ 3.0 Ma,晚中新世期间从南美到北美的早期迁移时间为~ 9 Ma,从北美到南美的早期迁移时间为~ 7 Ma(例如,Carrillo et al., 2015)。抵达南美洲的新北极有蹄类动物包括骆驼类、鹿类、蹄类、马科、貘类和gomphotherids(拟鼻目)(Webb, 2006)。在阿根廷3.7 Ma(根据Cione et al., 2007年,Chapadmalalan年龄)沉积物中发现的一种peccary (Tayassuidae)被认为是最早到达南美洲的偶蹄动物之一。在亚马逊盆地中新世(~ 10 Ma)的沉积物中也发现了Tayassuids、dromomerycine偶蹄动物、貘和gomphothae (Frailey & Campbell, 2012);尽管其中一些分类群的分类和年龄受到质疑(Gasparini et al., 2021;mother & Avilla, 2015)。
在这篇文章中,我们回顾并描述了圣地亚哥·罗斯于19世纪在阿根廷潘潘地区更新世沉积物中收集的新北极有蹄类化石(gomphotherids, camelids, tayassuids, cerids和equids),这些化石目前保存在苏黎世大学(最初在Roth, 1889年的目录5中)和日内瓦自然历史博物馆(未知目录;Voglino et al., 2023),瑞士。阿根廷的潘潘地区被解释为巴塔哥尼亚干旱地区和巴西湿润地区之间的过渡区(García-Morato等人,2021)。
罗斯对阿根廷地质学和古生物学的贡献是显著和众多的,其中包括化石收集和潘潘类哺乳动物协会的研究。他的实地工作伴随着一系列不同的出版物(Fernández, 1925;Giacchino & Gurovich, 2001;Sánchez-Villagra等,2023;Voglino, 2020;Voglino et al., 2023)。尽管Roth带到瑞士的化石是130多年前在阿根廷潘帕斯地区收集的,但对这一历史收藏品的研究仍然与分类学、古生物学和古生物地理目的有关,以及其他旨在了解更新世期间该地区哺乳动物动物群古多样性的研究(见Lynch et al., 2020;Le Verger, 2023;Ruiz-Ramoni et al., 2023;Voglino et al., 2023)。
这里研究的化石收藏包括共245种gomphotherids, camelids, tayassuids, cerids和马科动物的颅骨和颅骨后遗骸(附加文件1)。这些标本被存放在苏黎世大学(PIMUZ a /V)的古生物收藏馆和日内瓦自然历史博物馆地质和古生物学系(MHNG GEPI V-)。PIMUZ的标本对应于Roth(1889)出版的目录5;见附加文件1)。MHNG GEPI的标本与Roth (Voglino et al., 2023)出版的任何目录都没有关联(如果它们更早,我们不知道)。
分类学鉴定包括广泛的书目审查和对保存在瑞士这两个机构的化石和现存标本的比较:CFA, Colección Mastozoológica, Fundación de Historia Natural ' f
lix de Azara ',阿根廷布宜诺斯艾利斯;美洲农业科学院、联合国人口基金和人口基金、调查中心Antropológicas、Arqueológicas y Paleontológicas委内瑞拉弗朗西斯科·德米兰达实验大学;IGM, Museo Geológico joseroyGómez, Servicio Geológico columbiano,波哥大
,哥伦比亚;MACN,阿根廷自然科学博物馆“Bernardino Rivadavia”,布宜诺斯艾利斯,阿根廷;委内瑞拉苏利亚大学博物馆Biología MBLUZ;MCNC,加拉加斯博物馆,委内瑞拉;圣达菲省自然科学博物馆“弗洛伦蒂诺·阿梅吉诺”艺术硕士;MUFYCA,弗洛伦蒂诺大学博物馆,Carlos Ameghino,罗萨里奥,圣达菲;MLP,拉普拉塔博物馆,阿根廷;法国巴黎国立自然历史博物馆MNHN;瑞士巴塞尔自然历史博物馆NMB。
上前磨牙(P),下前磨牙(P),乳牙(dP/ dP),上磨牙(M),下磨牙(M),上门牙(I),下门牙(I),掌骨(MC),跖骨(MT)。
这里研究的标本来自位于布宜诺斯艾利斯省的19个化石遗址(附加文件1),罗斯(1889)将这些遗址称为:阿罗约·杜尔塞、阿罗约·佩加米诺、拉马洛、巴拉德罗、托莱内罗、Sánchez、圣Nicolás、蓬塔·圣埃琳娜、圣佩德罗和该省不确定的地区,以及圣达菲:阿罗约Pavón、圣洛伦佐、维拉Constitución、Río Carcara?á和罗萨里奥。上面提到的一些地方被概括了,它们的名字被缩短了,以优化图1中地图上的空间和地名(见附加文件1)。地点“Arroyo del Medio”位于布宜诺斯艾利斯和圣达菲省之间的边界,但确切位置未知。在目录5中,很少有标本没有指定的地点(见附加文件1)。
图1
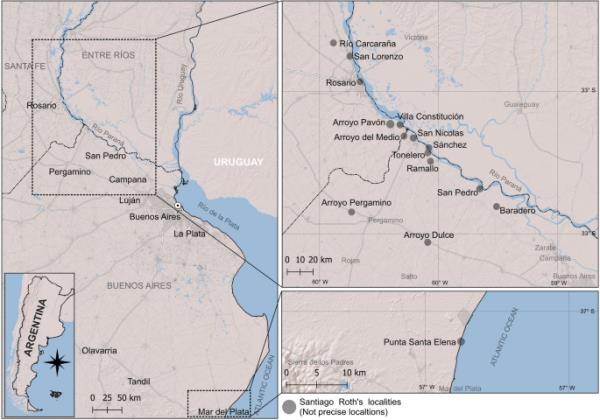
圣地亚哥·罗斯在目录5中提到的潘潘亚化石分布图(罗斯,1889年)。地点并不准确。该地图使用QGIS V.3 3.22.0制作,形状来自阿根廷Geográfico国家研究所(www.ign.gob.ar)。
Roth(1889)认为目录5中的化石来自“pampsamen infacrieur”、“intermsamen /Moyen”和“supsamrieur”,根据哺乳动物化石定义的生物带,这些化石大致相当于更新世哺乳动物阶段/时代的沉积物,分别是上Ensenadan-Bonaerian和Lujanian (Cione & Tonni, 2001;Cione et al., 2015)。每个标本的位置和地层来源(根据Roth, 1889年)都在附加文件1中提及。关于这里提到的地方的地质和地层学的更详细信息,见Voglino等人(2023)。
摘要
Resumen
介绍
材料与方法
系统的古生物学
有限公司
ncluding讲话
数据和材料的可用性
参考文献
致谢
作者信息
道德声明
补充信息
相关的内容
搜索
导航
#####
伊利格(1811年
gomphotheiidae Hay, 1922
Notiomastodon Cabrera, 1929a
北大乳齿象(Ameghino, 1888a)。
(图2 (a1b2)。
图2
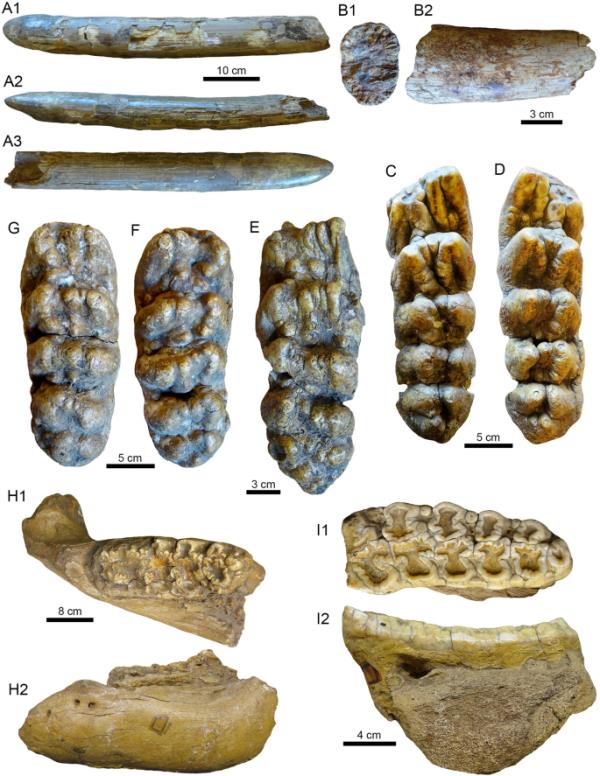
潘潘地区的Gomphotherids颅骨残骸。(a1b2。Notiomastodon platensis。a1a3。?右上方任务(MHNG GEPI V-3476)。b1 b2。上牙碎片(MHNG GEPI V-3504)。C-I2。北大乳齿象。c - d。左(C: MHNG GEPI V-3475)和右(D: MHNG GEPI V-3473) m3来自同一个人。E.分离左m3 (MHNG GEPI V-3478)。做减法。同一个体的左(G)和右(F) M3 (MHNG GEPI V-3474)。H1-H2。带有m3 (PIMUZ A/V 4092)的左半下颚碎片。I1-I2。左m3 (PIMUZ A/V 4161)。视图:横切面(B1),背侧(A2, H1),唇侧(I2),左侧侧边(H2),侧边(A1, A3, B2),咬合(C-G, I1)
两个上牙碎片(MHNG GEPI V-3476和MHNG GEPI V-3504)。标本的地理数据见附加文件1。
MHNG GEPI V-3476上牙为直角、完整、短直、不扭曲,长650mm,截面椭圆形,最大直径217mm(图2A1-A3)。没有观察到牙釉质层或带,沿象牙表面存在平行褶皱(Alberdi & Prado, 2022;moth
等人,2013)。该标本的远端呈现出明显的磨损,这是长鼻猿刮擦树木和灌木丛的特征(例如,Semprebon等人,2022)。较小的象牙碎片MHNG GEPI V-3504长度约为160 mm,也是直的,截面为椭圆形,最大直径为202 mm(图2B1-B2)。这块碎片属于象牙的最末端,因为没有可见的根的痕迹,也没有观察到牙釉质层或带。
一个多世纪以来,南美洲长鼻目动物的分类学在属和种的总数上一直存在长期而有争议的争论(Alberdi et al., 2002;Alberdi & Prado, 2022;Campbell et al., 2009;moth
等人,2012,2013,2017;mother & Avilla, 2015;卢卡斯,2013;普拉多等人,2005,以及其中的参考文献)。近几十年来,人们达成了共识,认为南方大陆上只有两个齿形兽属是正确的,一个是分布于安第斯山脉国家的Cuvieronius hyodon,另一个是分布于大陆从沿海到山区的Notiomastodon platensis (=Stegomastodon platensis) (Alberdi & Prado, 2022;moth
等人,2013,2017;mother & Avilla, 2015)。主要的颅骨特征被用来区分这两个属,而象牙对这些比较是有价值的。Cuvieronius的上牙细长,横截面呈亚圆形,在直到略上弯之间变化,明显扭曲,沿牙体有典型的珐琅带(moth
等人,2017a);卢卡斯,2013;母亲等人,2016)。相比之下,Notiomastodon的特征是上部象牙在长度、弯曲度和坚固度(横截面为圆形到椭圆形)上有很大的形态差异,但它们从不扭曲,在幼兽和一些成年个体中,牙釉质可能缺失或以单带形式存在或覆盖整个象牙(moth
等人,2017a)。根据母亲等人(2017a)的研究,在Notiomastodon中,形成上部象牙内部结构的牙本质锥体之间没有扭转,这是MHNG GEPI V-3476和MHNG GEPI V-3504的情况。这里描述的象牙的特征,包括直线形状,椭圆形截面,缺乏珐琅质带/层,与platensis上部象牙的特征一致。
北大乳齿象。
(图2C-I2, 3A-D)。
图3
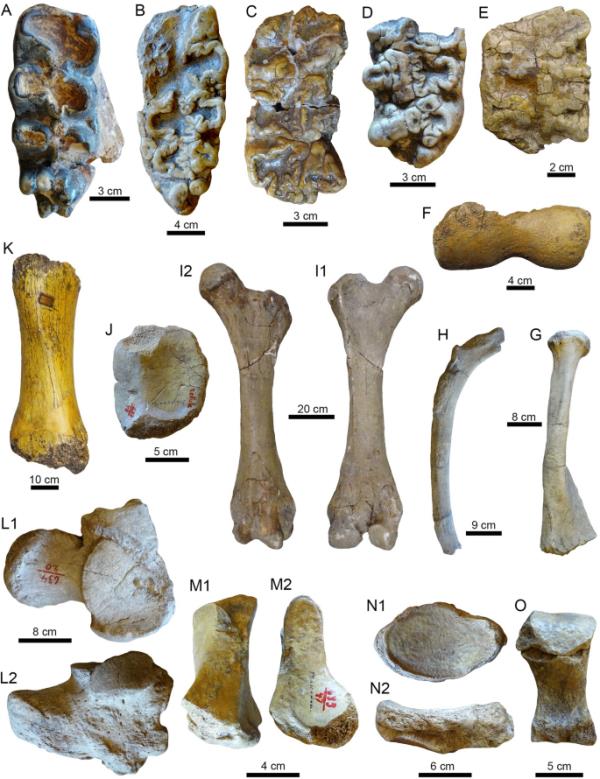
潘潘地区的Gomphotherids颅骨和后颅骨残骸。模拟。北大乳齿象;右m3 (A: MHNG GEPI V-3479),左m3 (B: MHNG GEPI V-3502),左M1 (C: MHNG GEPI V-3491),右M1 (D: MHNG GEPI V-3480)。开头的。嵌齿象科indet。E.不确定位置Molariform (PIMUZ A/ v4136)。F.左肱骨远端骨骺(PIMUZ A/V 4159)。G.左半径(MHNG GEPI V-3477)。H.右撕裂片段(MHNG GEPI V-3499)。I1-I2。左股骨(PIMUZ A/V 4158)。右髌骨(MHNG GEPI V-3490)。K.右胫骨不全(PIMUZ A/V 4160)。寓于“。右钙质(mhngepi V-3505)。M1-O。右梯形(M1-M2),左舟骨(N1-N2)和跖骨III (O) (MHNG GEPI V-3494)。视图:前部(K, I2, F-H, N2),背部(L1, N1),右侧(L2),侧面(M1),咬合(A-E),足底(O),后部(J, I1),腹侧(M2)
由于磨损阶段非常相似(MHNG GEPI V-3473和MHNG GEPI V-3475,图2C-D),可能属于同一个体的下三磨牙,根部破碎,第一和第二磨牙轻微磨损,左下第三磨牙破碎,无磨损,无牙根(MHNG GEPI V-3478,图2E),可能属于同一个体的两个上三磨牙,根部破碎,无磨损(MHNG GEPI V-3474,图2F-G)。左下颌分支碎片带第三磨牙,磨损严重(PIMUZ a /V 4092,图2H1-H2),左下第三磨牙,磨损严重,下颌根部和碎片(PIMUZ a /V 4161,图2 1 - 2),右下第三磨牙,中度磨损,第三磨牙碎片(MHNG GEPI V-3479,图3A),左下第三磨牙,中度磨损,舌前边界碎片(MHNG GEPI V-3502,图3B)。左上第一磨牙破碎,中度磨损(MHNG GEPI V-3491,图3C和-3486),右下第一磨牙破碎,第一磨牙缺失部分,中度磨损(MHNG GEPI V-3480)。标本的地理数据见附加文件1。
所有臼齿均为平齿,具有平齿Notiomastodon platensis特有的双至简单三叶草磨损模式(即MHNG GEPI V-3502, -3491, -3486和-3480)。第三磨牙为五突齿,远端带不固定,无磨损或中度磨损的牙齿为57 - 64杯,在文献中描述的Notiomastodon的35 - 82杯范围内(moth
等人,2017c)。完整臼齿的总长度见附加文件1。
分离的臼齿是南美洲长鼻目动物中最常见的化石,但不幸的是,当分离和/或碎片化时,它们并不能完全诊断Notiomastodon和Cuvieronius,因为大多数特征是可变的和重叠的(moth
等人,2017b)。完整且保存完好的第三磨牙的尖牙总数已成功用于区分这两个属(moth
& Avilla, 2015;母亲等人,2017c)。Cuvieronius hyodon的第三磨牙有33到60个尖牙,而Notiomastodon的第三磨牙有35到82个尖牙(见moth
等人,2017c)。如上所述,我们在这里描述的完整的第三磨牙显示出57到64个尖牙的范围,属于Notiomastodon的范围。一些标本的碎片性质不允许获得尖的完整数量,但由于前和后的磨损模式是复杂的(双三叶草),这表明一些标本的尖的数量可能更高(例如,PIMUZ A/V 4092, 4161, MHNG GEPI V-3502)。本文报告的标本暂定为平齿Notiomastodon cf. N. platensis。
嵌齿象科indet。
(图3开头的)。
5颗严重磨损的断牙和不全牙(PIMUZ A/V 4105、4136、4185、4279)、左肱骨远端骨骺(PIMUZ A/V 4159)、左桡骨(MHNG GEPI V-3477)、左股骨(PIMUZ A/V 4158)、右髌骨(MHNG GEPI V-3490)、右胫骨(PIMUZ A/V 4160)、右跟骨(MHNG GEPI V-3505)、舟骨、右椎体、跖骨III和掌骨III (MHNG GEPI V-3494)和肋骨碎片(MHNG GEPI V-3499)。标本的地理数据见附加文件1。
PIMUZ A/V 4105是一颗半下颌骨碎片,牙冠不完整和破碎,牙齿碎片不确定,保存状态较差。编号PIMUZ A/V 4279对应于一个不完整且磨损严重的磨牙,可能是M2或M2,以及另外两个位置不确定的磨牙状碎片。PIMUZ A/V 4185由三个碎片和孤立的lophs代表,可能来自同一个体。PIMUZ A/V 4136是一个位置不确定的极端磨损的破碎磨牙(图3E)。Postcranial附属物的元素是由一个左的肱骨远端骺(PIMUZ a / 4159 V,图3 f),长度567毫米半径失踪近端骺(MHNG罗格·V - 3477,图3 g),完成左股骨的总长度1045毫米(PIMUZ a / 4158 V,图3 i1-i2),右胫骨骨干的445毫米的长度(PIMUZ a / 4160 V,图3 k), a ?右膝盖骨(MHNG罗格·V - 3490,图3 j),一个完整的右跟骨(MHNG罗格·V - 3505,图3寓于),舟状,梯形,跖骨III和掌骨III (mhngepi V-3494,图3M1-O)。破碎的右肋骨是轴向骨架的唯一元件(MHNG GEPI V-3499,图3H)。
牙齿遗骸的保存状态以及对颅骨后骨骼缺乏诊断特征(moth
& Avilla, 2015),不允许对这些标本进行超越家庭水平的分类。Cuvieronius和Notiomastodon的颅后特征没有详细的形态学研究可以证明颅后特征的分类学意义。考虑到已知的Notiomastodon的地理分布,包括Pampean地区(Alberdi & Prado, 2022;卢卡斯,2013;mother & Avilla, 2015;母亲等人,2017a, b, c),最终可能分配到这个分类单元。
偶蹄动物(欧文,1848)。
葛雷(1821年
喇嘛居维叶(1800年
喇嘛·瓜内科(米
勒,1776年)。
(图4 a1a3)。
图4

潘潘地区的骆驼类头骨残骸。a1a3。藏獒的下颌骨(PIMUZ A/V 4089)。B1-B6。左上颌骨(B1-B2),颅骨前部(B3)和左下颌骨(PIMUZ A/V 4287)。C1-G2。半可食用的Hemiauchenia cf. H. paradoxa片段(右:C1-C2: PIMUZ A/V 4186, D1-D2: PIMUZ A/V 4187, E1-E2: PIMUZ A/V 4192, F1-F2: PIMUZ A/V 4257;左:G1-G2: PIMUZ A/V 4195)。视图:背侧(A2, B4),左侧(B1, B5, G2),舌侧(C2, F2),右侧(A1, B3, D2, E2),咬合(A3, B6, C1, D1, E1, F1, G1)和腹侧(B2)。犬齿(c)、乳牙前磨牙(dp)、牙窝(fos)、下牙窝(hyd)、门牙(i)、下磨牙(m)、下前磨牙(p)、副柱盖(pald)、前磨牙牙槽(ap)、原柱盖(prld)、上磨牙(m)、上前磨牙(p)
几乎完整的下颌骨,下磨牙系列不完整,没有冠状突、角突和髁突(PIMUZ A/V 4089)。标本的地理数据见附加文件1。
下颌骨纤细,在冠突和角突的水平处断裂。水平支的背缘和腹缘几乎彼此平行。水平支舌面平,唇侧微凸。由于骨表面嵌有沉积物,未观察到吻侧和尾侧的颏孔。有五个门牙。犬科动物没有保存下来。牙式显示,在Lama guanicoe和Hemiauchenia paradoxa中,前磨牙的牙槽位于第三和第四位置(Lynch et al., 2020)。右半下颌骨有m1-m3,而左侧保留m1-m2系列,在m3的前根水平断裂(图4A2)。咬合长度和宽度m1分别为13 mm和10 mm, m2分别为18 mm和11 mm, m3分别为23 mm和10 mm。右侧m1 ~ m3系列长度为54mm。这些月齿臼齿的特征是具有发育良好的前柱头或“羊驼背”(原柱头和旁柱头),尽管在m3上这些是弱突出的阴唇-舌。m1上的前窝(trigonid fossa, Scherer et al., 2007)由于磨损程度较深而未见,因此该标本对应Breyer(1977)分类中“磨损阶段4”的成人。所有的窝都是闭合的,这是Camelidae比Cervidae更典型的特征(后者的前窝远端舌打开)(图4A3)。唇形为u形(例如,Gasparini et al., 2017;Scherer et al., 2007),并且每个磨牙的第二叶通过明显的垂直和深唇弯曲与前叶分开。第三叶(下圆锥)较小,轮廓呈椭圆形(图4A3)。
PIMUZ A/V 4089显示出许多与南美Camelidae一致的形态特征(见Lynch et al., 2020;谢勒,2013)。上述形态学和形态计量学特征允许我们将PIMUZ A/V 4089分配给喇嘛瓜。Lama的化石记录在阿根廷、玻利维亚、巴西、智利、哥伦比亚、厄瓜多尔、秘鲁和乌拉圭的晚更新世沉积物中被发现(Scherer, 2013;Tonni & Politis, 1980,以及其中的参考文献)。然而,根据Wheeler(1995),来自厄瓜多尔和哥伦比亚的Lama的记录需要确认。Lama在阿根廷南部地区的全新世沉积物中得到了广泛的记录,尽管在这段时间内,在帕拉那河流域中下游不同地点的记录受到了质疑(见Politis et al., 2011)。
Menegaz和Ortiz Jaureguizar(1995)报道了在北潘帕(布宜诺斯艾利斯省)更新世沉积物中发现的一种从解剖学上与L. guanicoe无法区分的Lama化石的存在,其尺寸与Hemiauchenia的范围相似。Lynch等人(2020)报道了阿根廷早更新世保存完好的南美Lamini部分骨骼,并对茶树亚科进行了新的分支分析。该标本(PIMUZ A/V 4165)也是由Santiago Roth在San Nicolás地区(布宜诺斯艾利斯省)的paran
河右侧悬崖上收集的(Roth, 1889)。根据Lynch等人(2020)假设的系统发育位置,PIMUZ A/V 4165与L. guanicoe和Vicugna Vicugna的亲缘关系比与H. paradoxa的亲缘关系更密切。另一方面,对Lama分类学的形态学、形态测量学和一些分子方面的考虑,认为已灭绝的Lama gracilis是巴塔哥尼亚和火地岛晚更新世-全新世早期的一种细毛虫vicu?a的有效Lamini分类群(Menegaz, 2000;Menegaz et al., 1989;Metcalf et al., 2016)。然而,关于南美洲现存和化石骆驼科动物的分类地位和系统发育背景尚未达成共识(见Cajal et al., 2010;Cartajena et al., 2010;米勒,1924;Stanley et al., 1994;Weinstock et al., 2009)。因此,这些假设必须通过更多的分子、形态学和形态计量学研究来检验。
目前,已知的野生Lamini Camelidae有两种:L. guanicoe和V. vicugna,它们局限于安第斯山脉和巴塔哥尼亚山脉中北部地区(8oS至55oS)的干旱至半干旱环境,以及海拔高度从海平面到4000 m之间(Baldi et al., 2008;富兰克林,1982;Lichtenstein et al., 2008;惠勒,1995)。
Hemiauchenia Gervais & Ameghino, 1880
Gervais & Ameghino, 1880
(图4 b1-b6)。
颅骨前区和左上颌骨有P3-M3,左半下颌骨有三对下门牙和P3-M3,来自同一个体(PIMUZ A/V 4287)。标本的地理数据见附加文件1。
颅骨残骸包括破碎的前颌骨和部分保存至眶下孔区域的上颌骨,右I3和犬形(图4B3)。左上颌骨为P3-M3系列,有轻微磨损,M3出牙。M3中的两个化石都被打开(图4B2)。磨牙串咬合长度为82毫米。PIMUZ A/V 4287对应于一个年轻的成年骆驼(Breyer, 1977)。左半下颌骨两侧有三个下门牙(i1-i3),一个犬齿(通过隔膜与门牙分开)和p3-m3系列(图4B-B6)。磨牙串咬合长度为105mm。水平支舌面平坦,唇侧微凸(图4B4)。在下颌骨的唇面上可以观察到嘴侧和尾侧的颏孔。前者位于犬科动物的远端(图4B5)。左犬形被保存下来,两者都是m3。p3-m2系列唇形唇突闭合(Gasparini et al., 2017;Scherer et al., 2007)。每颗颊齿的第二叶由深唇瓣与前叶分开。第三叶(下圆锥)较小,轮廓呈椭圆形。下磨牙的前柱体(原柱体和旁柱体)不可见,因为部分牙齿在牙槽内。除m3外,所有牙窝均闭合,显示两个牙窝均向中远端打开。PIMUZ A/V 4287对应的是一个年轻的成年个体,恒牙已经长出,但几乎没有磨损(Breyer, 1977)。
Hemiauchenia是上新世晚期至更新世期间居住在南美洲的最大的Lamini骆驼科动物之一(Gasparini et al., 2017;谢勒,2013)。主要从阿根廷和乌拉圭的露头岩层可知(Cione & Tonni, 1999;Cione et al., 1999,2015;Gasparini et al., 2017;Menegaz & Ortiz-Jaureguizar, 1995;Ubilla, 2004;Ubilla & Perea, 1999)。相比之下,在玻利维亚早更新世沉积层中发现了古olama (MacFadden & Shockey, 1997;Marshall et al., 1984),以及玻利维亚、巴西、智利、厄瓜多尔、巴拉圭、秘鲁和委内瑞拉的晚更新世矿床(Carrillo-Brice?o, 2015;cartellle et al., 1989;Marshall et al., 1984;谢勒,2013)。
López-Aranguren(1930)将H. paradoxa与Hemiauchenia major同义,Cabrera(1932、1935)和Rusconi(1931)将前者归为Palaeolama weddelli。后来的研究再次证实了悖论H.,虽然包括在另一个属:古螈(见Hoffstetter, 1952)。Webb(1974)和Scherer(2013)认为Hemiauchenia和Palaeolama都是有效的分类群,但认为H. paradoxa仅限于阿根廷,H. major仅限于巴西东北部。然而,一些作者认为巴西的形式对应于大古火山(Bergqvist, 1993;cartellle et al., 1989;gusamrin等人,1990;Souza-Cunha, 1966)。一些分类学研究表明,在形态上的差异并不能支持类人猿和大类人猿之间的分类学分离,而是表明种内变异(Cartelle, 1992,1999)。然而,最近Scherer(2009, 2013)支持了这两个属在南美洲的分类学有效性,并承认了至少三个古古马物种:P. weddelli, P. major和P. hoffsteteteri。相反,H. paradoxa被认为是该属的唯一有效种。
偏忆症。
(图4 c1-g2)。
2个带有乳牙的右半下颌骨碎片(PIMUZ A/V 4186),右半下颌骨部分保存m1-m3,左半下颌骨部分保存dp4-m2 (PIMUZ A/V 4187),右半下颌骨部分保存p3-p4 (PIMUZ A/V 4192),左半下颌骨部分保存taltalid m2和整个m3 (PIMUZ A/V 4195),右半下颌骨部分保存m2 - m3 (PIMUZ A/V 4201)。左、右半下颌骨含乳牙和恒牙2块(PIMUZ A/V 4200),右半下颌骨含m1-m3 (PIMUZ A/V 4257),离体m2 (PIMUZ A/V 4127, PIMUZ A/V 4179, A/V 4189),离体下牙(PIMUZ A/V 4196, PIMUZ A/V 4255)。标本的地理数据见附加文件1。
PIMUZ A/V 4186对应两个右下颚,dp4无磨损,m1喷发。dp4类似于臼齿系列的齿,有三个裂片。尖头仍然分开,没有磨损。化石向中远端打开(图4C1)。PIMUZ A/V 4187由两块可能来自同一个体的下颌骨碎片代表。右半下颌骨部分保留了m1-m3的爪骨(图4D1-D2)。左半下颌骨有一个dp4和两个m1-m2的三角体断裂并被碳酸盐结块覆盖。磨牙窝全部闭合,唇唇瓣呈u形。左dp4类似于molariform系列,有三个裂片。右图m3显示了典型的“羊驼扶壁”。PIMUZ A/V 4192是具有p3-p4系列的右半可下咽体片段(图4E1-E2)。p3是横向压缩的,只由一个叶组成;这颗牙没有磨损。p4有轻微磨损,在唇侧和舌侧都有弹性,形成两个裂片。中叶向中远端拉长,远端横向宽。
PIMUZ A/V 4195为左半下颚体,其爪形为m2和一个m3(图4G1-G2)。两磨牙窝闭合,m3上柱头发育。下圆锥较小,呈卵圆形,缺少窝。PIMUZ A/V 4200代表了带有牙齿的半下颚动物的左右碎片。右边的级数有根m1和完整的m2-m3。m3没有磨损,没有下圆锥,有一个小的原柱体,两个化石都是中向打开的。左边的级数是dp4-m2。PIMUZ A/V 4201是右半下颚的碎片,保留了m2在磨损后期的爪骨和整个m3。在m3中未见中柱,而下圆锥较小,轮廓为椭圆形,缺乏窝。PIMUZ A/V 4179,和PIMUZ A/V 4189, 4127是隔离的m2,第一个在左边,第二个和第三个在右边。牙齿有不同的磨损阶段,牙窝闭合,中柱发达。PIMUZ A/V 4179显示出“磨损阶段1”,其化石呈舌状打开,表明该标本属于幼年骆驼(Breyer, 1977)。PIMUZ A/V 4255对应于m1, m2和m3。牙齿不能保护牙根。PIMUZ A/V 4196由一个左永久的p4表示,可能还有p3, m1和m2。这些牙齿类似于先前在H. paradoxa (PIMUZ A/V 4287;见上文)。PIMUZ A/V 4257是具有m1-m3的右半球形(图4F1-F2)。牙齿有闭合的窝窝,尽管m3上的窝窝是突出的和中远端打开的。舌柱体(原柱体)存在于所有磨牙中,舌上发育良好,而附柱体仅在m1上保存,在m2上更为明显(图4F1)。
在南美骆驼的下乳牙中未观察到诊断特征(例如,Cabrera, 1932, 1935;Rusconi, 1931;韦伯,1974)。上述标本暂时归属于Hemiauchenia cf. H. paradoxa,因为它们的大小和牙齿形态与阿根廷和巴西更新世沉积物中的H. paradoxa相似(Scherer et al., 2007;Vezzosi et al., 2019)。
参:赤霉病属。
(图5 a1-e2)。
图5
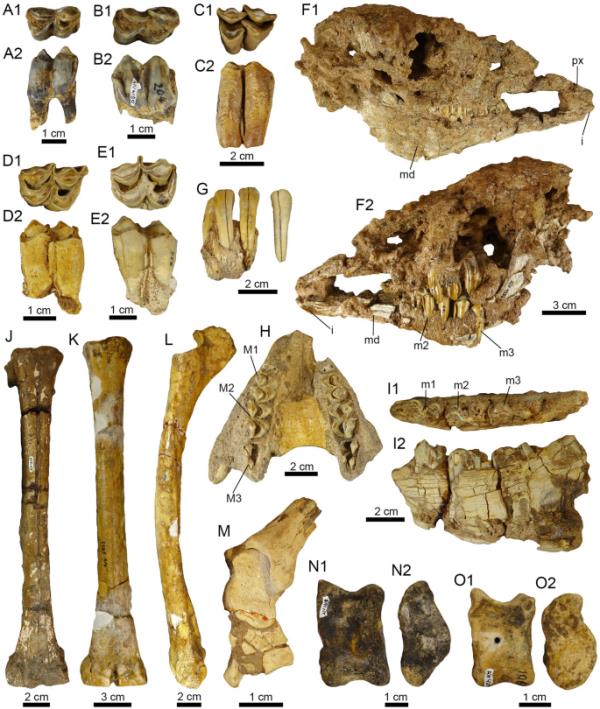
潘潘地区的骆驼头盖骨和后头盖骨。A1-E2。cf. Hemiauchenia sp. (A1-B2: PIMUZ A/ v4205;C1-c2: a / v 4182;D1-d2: a / v 4098;E1-e2: pimuz a / v 4181)。F1-O2。骆驼科不定。F1-F2。幼年骆驼的头骨和下颌骨(PIMUZ a /V 4286)。G.带有四门牙的上颌骨碎片(PIMUZ A/V 4253)。H.两个前上颌骨(PIMUZ A/V 4256)。I1-I2。带有牙列的上颌骨碎片(PIMUZ A/V 4253)。J.左掌骨(PIMUZ A/V 4250)。K-L。右掌骨(K)和右桡骨-尺骨(L) (PIMUZ A/V 5969)。M.左胫骨远端有黄芪、舟状骨和长方体(PIMUZ A/V 4090)。N1-O2。左黄芪(PIMUZ A/V 4208)。视图:前(G),背(J-K, N1, O1),唇(A2, C2),舌(B2, D2, E2),内侧(L, N2, O2),咬合(A1, B1, C1, D1, E1, I1),左侧(F2, I2, M),右侧(F1)和腹侧(H)。门牙(i),下磨牙(M),下颌骨(md),前颌(px)和上磨牙(M)
孤立的臼齿可能是M1?或平方米?(pimuz a / v 4098, 4103, 4180, 4181, 4182)。标本的地理数据见附加文件1。
材料由四个左边(PIMUZ A/V 4103、4180、4182)和一个右边(PIMUZ A/V 4181)表示,可能是M1或M2,具有不同磨损阶段的特征(图5A1-E2)。中叶比远叶大,有非常大的花柱(亚柱、副柱和中柱),三角窝较深。
这些臼齿的大小与Hemiauchenia (Cabrera, 1935;Scherer et al., 2007),舌纹呈u形,与大古兽的v形牙齿舌纹不同(见Scherer et al., 2007;韦伯,1974)。根据一项基于北美标本的研究,lophs的形状是Hemiauchenia和Palaeolama之间的一般差异(Webb, 1974)。需要更多的南美骆驼样本来测试在上牙列中观察到的其他牙齿特征之间的这种普遍差异。
骆驼科indet。
(图5 f1-o2)。
遗骨包括4个下门牙,上颌和右半下颌骨碎片,均有牙列(PIMUZ A/V 4253),保存较差的幼年骆驼头骨,下颌骨和牙列(PIMUZ A/V 4286),下颌骨和前上颌碎片(PIMUZ A/V 4256), 75个分离的臼齿(PIMUZ A/V 4254),分离和碎片的半下颌骨,共同骨和6个门牙和臼齿牙齿(PIMUZ A/V s/n),没有地理背景;颅后遗骸包括左侵蚀掌骨(PIMUZ a /V 4250),左胫骨远端黄骨,舟骨和长方体(PIMUZ a /V 4090),右桡骨-尺骨和右掌骨,可能来自同一个人(PIMUZ a /V 5969),三个侵蚀的黄骨,一个舟骨和一个足跖指骨(PIMUZ a /V 4208)。标本的地理数据见附加文件1。
PIMUZ A/V 4253的左侧上颌骨碎片为M1 - m3系列,M1完全被碳酸盐结块覆盖。下牙列有右i1-i2和左i1-i2(最后一个门牙是分离的)(图5G)。右侧半下颌体p4和m1-m2远端根出窝,m3出窝(图511 - 2)。所有的臼齿都与大型骆驼的臼齿大小相似,尽管咬合表面断裂或被碳酸盐覆盖,使它们无法进行比较。皮姆兹A/V 4286头骨呈现出一种名为喇嘛的小型骆驼的形态和尺寸。然而,粘附在该标本上的碳酸盐使诊断特征无法识别。上颌骨和下颌骨在这个标本中没有提供诊断元素的一般分配。PIMUZ A/V 4254由许多不同大小的Camelidae牙齿(N°75)代表,其保存状态不允许更准确的测定。编号为PIMUZ A/V 4250(图5J)的左掌骨III和IV彼此融合,除了远端,如在骆驼类中所见。PIMUZ A/V 4090表现为左胫骨远端与黄体、舟状骨和长方体关节相连;碳酸盐的覆盖层(图5M)阻碍了超出家庭水平的分配。PIMUZ A/V 5969是部分受损的右尺骨桡骨(图5L)和右掌骨(图5K)。PIMUZ A/V 4208由三个侵蚀的黄芪(图5N1-O2),一个舟状指骨和一个足状指骨代表。三种黄芪中有两种在大小上与小型骆驼属(如Lama guanicoe)相似,而较大的黄芪的形态和大小与Hemiauchenia相似(Scherer et al., 2007)。舟骨指骨和足骨指骨都不允许更精确的测定。
由于保存不良和/或缺乏诊断特征,无法对上述标本进行具体定位。
帕尔默(1897年
塔亚苏·费舍尔·冯·瓦尔德海姆(1814年
塔亚苏pecari(链接,1795年)。
(图6 a1a2)。
图6

潘潘亚地区的塔亚suidae遗迹。a1 a2。左半可食用的碎片,m2和m3的根基本完整(PIMUZ A/V 4188)。b1 b2。左/右?落叶p4 (PIMUZ A/V 5258)。视图:唇/舌(B2),舌(A2),咬合(A1, B1)。中央圆锥(ccd)、牙状圆锥(dn)、内圆锥(ec)、内圆锥(entd)、下圆锥(hd)、下圆锥(hyd)、外侧圆锥(lacd)、舌状圆锥(licd)、后圆锥(mtcd)、后圆锥(pcd)和原圆锥(prd)
左半可食用的碎片,m2和m3的根基本完整(PIMUZ A/V 4188)。标本的地理数据见附加文件1。
PIMUZ A/V 4188是左半可食的碎片,m2的末根和m3的基本完整。牙齿为短齿和布齿,保留了三个主要的尖齿,有轻微的磨损:中侧的后突,与下突之间有一个山谷隔开,远端是内突(图6A1)。磨牙在原锥体的水平处断裂,因此我们无法检查有圆齿的齿环的存在(例如,Gasparini, 2007)。第一对前尖(后圆锥和原圆锥)部分断裂,但两者加在一起比后一对尖(下圆锥和内圆锥)略宽,保存完好(图6A1)。在这两个叶的后面,有一个结构复杂的第三叶,其中可以看到四个不同大小的尖头,如塔亚苏pecari(见Gasparini, 2007;Parisi Dutra等人,2017)。继Gasparini(2007)和Parisi Dutra等人(2017)之后,PIMUZ A/V 4188属于T. pecari的大小范围,中远端长度为22 mm,舌唇宽度为12 mm。这个m3在中远端被拉长,由于第三个尖叶的存在而形成了一个复杂的下圆锥,因此具有近矩形的轮廓(Frailey & Campbell, 2012)。
继Gasparini(2007)和Parisi Dutra等人(2017)之后,这里报道的m3的结构明显不同于Brasilochoerus spp., Parachoerus wagneri, Tayassu tajacu和Platygonus spp.(除了Platygonus chapadmalensis),因为这些tayassuids有一个简单的第三叶(具有独特的优势尖;见Gasparini, 2007),有时伴随着一对较小的尖头(Frailey & Campbell, 2012;Gasparini & Ferrero, 2010)。
鼬科是GABI期间最早进入南美洲的哺乳动物群体之一(woodburn, 2010)。它们广泛的化石记录与阿根廷、巴西、乌拉圭、玻利维亚、哥伦比亚、秘鲁和委内瑞拉的晚新生代矿床有关(Gasparini, 2013;Montellano-Ballesteros et al., 2014;Parisi Dutra et al., 2017,以及其中的参考文献)。它首次到达的确切时间是有争议的,估计日期在3.7到3.1 Ma之间(见Cione等人,2007年,2015年;Woodburne, 2010)。根据化石记录,更新世南美洲的塔亚松类的多样性和丰度高于上新世和全新世(Gasparini, 2013)。此外,南美洲的塔亚suids化石物种多样性和丰度较高的地区集中在阿根廷(Gasparini, 2013),根据Gasparini(2007),在阿根廷发现了3属12种塔亚suidae。在更新世-全新世边界附近,鱼类的多样性显著减少(Gasparini, 2013),只有两个属(Parachoerus和Tayassu)和三个物种存活至今(Gasparini, 2013;Parisi Dutra等人,2017)。其中,白唇pecari广泛分布在新热带地区,从阿根廷北部到墨西哥东南部(Bustos等人,2019;Gasparini et al., 2014)。尽管其分布范围大,在南美洲有化石记录(Gasparini, 2013;Montellano-Ballesteros et al., 2014),其过去的发生很少,并且仅限于布宜诺斯艾利斯,Corrientes, Misiones, Santa Fe和Santiago del Estero的中更新世-全新世(Gasparini, 2013;Gasparini et al., 2014)。T. pecari的一些形态特征可能与它们所居住的潮湿气候条件和林地环境有关(Gasparini et al., 2014)。然而,塔亚苏pecari占据了广泛的地理范围,必须具有广泛的生态耐受性,这使得很难预测其生活的栖息地(Parisi Dutra et al., 2017;Bustos et al., 2019)。
Tayassuidae indet。
(图6 b1 b2)。
左/右?dp4至少有两个叶(PIMUZ A/V 5258)。标本的地理数据见附加文件1。
PIMUZ A/V 5258是一种落叶突齿和短突齿dp4,其叶片咬合面有较严重的磨损,表明该标本属于年轻的成年个体。在PIMUZ A/V 5258中,臼齿化明显发生在化石和现存的狐齿科的乳牙中(Frailey & Campbell, 2012;Gasparini, 2007;Gasparini et al., 2011)。由于磨损,PIMUZ A/V 5258的齿形(如主尖和副尖)的详细描述相当困难。
由于这颗离体牙齿(PIMUZ A/V 5258)保存不完整,是否有系统的分配值得怀疑。
Cervidae Goldfuss(1820年
莫雷内拉弗斯·卡莱特,1922年
Morenelaphus sp。
(图7 a1a3)。
图7
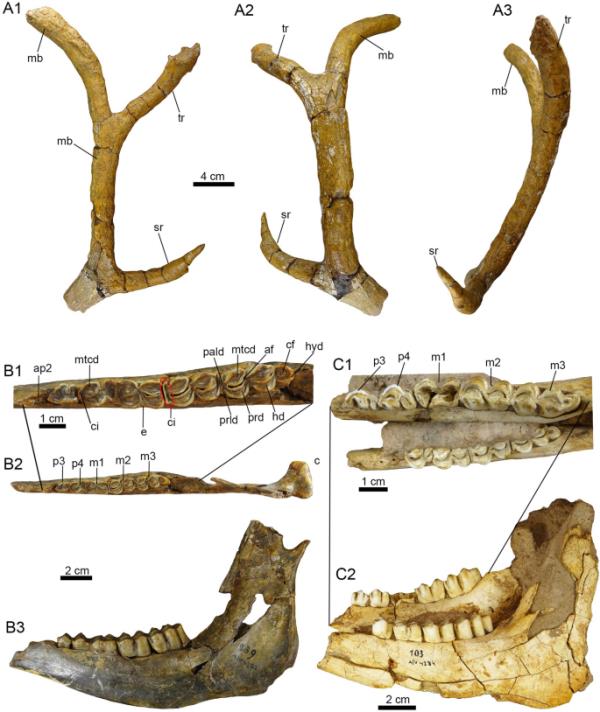
潘潘亚地区的Cervid遗迹。a1a3。Morenelaphus sp. (PIMUZ A/V 4162)左角不完整。B1-C2。不确定Cervidae的左下颌骨(B1-B3: PIMUZ A/V 4252)和下颌骨(C1-C2: PIMUZ A/V 4284)。视图:前部(A3),背部(B2),外侧(A2),左侧(B3, C2)内侧(A1),咬合(B1, C1)。前窝(af)、前窝(ci)、圆窝(cf)、外柱头(e)、下圆锥(hd)、下圆锥(hyd)、下磨牙(m)、下前磨牙(p)、平均梁(mb)、后圆锥(mtcd)、副柱头(pald)、前磨牙牙槽(ap)、原圆锥(prd)、原柱头(prld)、第二分枝(sr)、第三分枝(tr)
一个不完整的左鹿角(PIMUZ A/V 4162)。标本的地理数据见附加文件1。
PIMUZ A/V 4162由一个长而复杂的不完整的左角组成(图7A1-A3)。鹿角的保存部分显示平均梁,在前面的第二分枝附近破碎,具有圆形的横截面(Carette, 1922;Kraglievich, 1932)。此外,第二分支和第三分支是向前的,而平均光束是向后的。第二分支和第三分支在它们的轴线交点形成一个约45度的角。鹿角在其自身的轴线上扭曲,并在鹿角的扭转之后,沿其外表面有轻微的纵向凹槽。
根据Ameghino (1888b)和Alcaraz(2010)的研究,Morenelaphus的平均光束在第三次分叉后明显向后弯曲,正如在PIMUZ A/V 4162中观察到的那样。传统上,鹿角被用作南美鹿科化石中不同分类水平的诊断结构(Alcaraz, 2010;Ameghino, 1888 b;Kraglievich, 1932)。其他一些研究使用了其他结构用于分类目的(Orcesi等人,2019;Pego, 2014;韦2015;Vezzosi & Chimento, 2021)。PIMUZ A/V 4162是一个没有任何其他相关结构的鹿角碎片,它不允许分配超出Morenelaphus sp。
Morenelaphus以其长而巨大的鹿角而闻名(例如,Ameghino, 1888b;卡布瑞拉,1929 b;Carette, 1922)。在阿根廷潘潘地区的更新世沉积物中发现了两种Morenelaphus brachyceros和Morenelaphus lujanensis。这两个物种的系统区分仅由其鹿角的形态学支持(Alcaraz, 2010;Ameghino, 1888 b;卡布瑞拉,1929 b;Gervais & Ameghino, 1880;Menegaz, 2000);尽管一些头骨和牙齿特征有助于区分这些化石形式(Chimento等人,2019;Rotti等人,2021;Vezzosi & Chimento, 2021;Vezzosi et al., 2020)。化石记录表明,这两个Morenelaphus物种在更新世属于不同的生物年代学间隔(Alcaraz, 2010;Cassini et al., 2016)。Morenelaphus被认为是阿根廷、乌拉圭和巴西南部至北部潘潘亚低地的典型灭绝鹿(Cassini et al., 2016;Chimento et al., 2019;费列罗等人,2007;Kraglievich, 1932;Rotti等人,2021;Ubilla & Martínez, 2016;Vezzosi et al., 2019)。一些研究表明,草是Morenelaphus饮食中最常见的元素(Rotti等人,2018),尽管Tomassini等人(2020)提出了更多的浏览器喂养偏好。
鹿科indet。
(图7 b1-c2)。
左半下颌骨为p2和p3-m3系列(PIMUZ A/V 4252),下颌骨为左侧p4-m3系列和p2 - p3根,右列牙列为p3-m3系列(PIMUZ A/V 4284)。标本的地理数据见附加文件1。
PIMUZ A/V 4252是部分保存的左下颚,具有p3-m3系列和p2的肺泡(图7B1-B3)。牙齿没有显示出磨损的晚期(图7B1)。磨牙串咬合长度为61 mm。PIMUZ A/V 4284保留了这两种半食用食品(图7C1-C2)。左齿列有p4-m3级数和p2-p3的根,右齿列保留p3-m3级数,但其m1舌断。右侧m1-m3系列为51mm,左侧m1-m3系列为52mm。牙齿是单硒齿;大多数尖端是分化的,尽管有发达的cristid。两个下颌骨都很强壮,其狭口比任何现存的南美鹿都短。
PIMUZ A/V 4252和PIMUZ A/V 4284的牙列与Cervidae (Heckeberg, 2020)一样具有六颗牙齿的前磨牙系列,而Camelidae则有五颗牙齿(Lynch et al., 2020)。短隔膜的存在是一种形态学特征,被认为是Morenelaphus的诊断(Alcaraz, 2010;Chimento et al., 2019;Menegaz, 2000;Pego, 2014)。Pêgo(2014)报道,在Morenelaphus的磨牙中,存在原锥体和下锥体的唇部呈u形形态;而活的蛇尾纲(Blastocerus, Odocoileus, Ozotoceros, subbulo, Pudu和Hippocamelus)有一个v形切片。在两个标本PIMUZ A/V 4252和PIMUZ A/V 4284的牙列中观察到u形形状(图7B1, C1)。然而,尚不清楚莫伦尼aphus的下颌遗骸是否与有牙列和/或鹿角的头骨相关(Vezzosi, 2015;Vezzosi & Chimento, 2021)。此外,在PIMUZ a /V 4252和PIMUZ a /V 4284以及Chimento et al.(2019)提出的该属的其他标本中不可能识别出短狭缝。事实上,在活鹿个体发育过程中,裂口的长度是高度可变的,必须在南美洲物种之间的大型比较框架中进行测试。在PIMUZ A/V 4252和PIMUZ A/V 4284中,牙列在咬合角度上是近平行的,位于m2-m3水平,在半下颌骨上有更多的舌部投影,而在Camelidae (Hemiauchenia paradoxa, Lama guanicoe和paleolama major)中,颊磨牙与半下颌骨平行,与唇舌方向平行(Lynch et al., 2020;Scherer et al., 2007)。前窝骨在p4-m3的三角骨舌中部分打开(例如,图7B1)。在m1和m2中没有后骨的痕迹;然而,在m3中,这个化石是开口的。下圆锥大,呈圆形。有一个圆形窝窝(图7B1)。在南美洲,骆驼科的齿形较小,呈椭圆形/近椭圆形,没有窝。p4可见前扣带,但发育较弱。由于保存和磨损阶段,我们在m1中未见前扣带。元锥体是p4三角体中最大的杯状体。m3具有元柱体,具有明显的原柱体和附柱体,这是更新世南美洲Camelidae中不常见的特征(Scherer et al., 2007)。根据Heckeberg(2020)的说法,鹿科动物的下臼齿中有1到3个外柱体。这些都不高,并且在老年人中受到磨损的影响。图7B1中可见到m1的外柱体。我们不排除PIMUZ A/V 4252和PIMUZ A/V 4284可能属于Morenelaphus或另一种美洲鹿,因为我们的比较不允许提供更具体的确定。有必要在更广泛的比较和系统发育框架中对这些特征进行修订和测试,以确定与现存或已灭绝的南美鹿的亲缘关系。
欧文(1848年
格雷(1821年
希皮迪恩·欧文(1869年
《原理论》(隆德,1846)。
希皮底翁cf. H. principale
(图8 (a1b2)。
图8
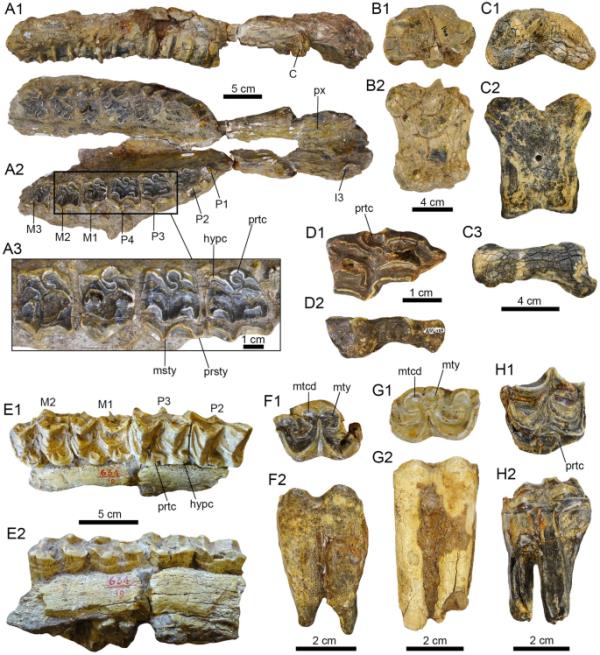
潘潘地区的马科动物。(a1b2。Hippidion cf. H. principale (PIMUZ A/V 4100);腹侧面颅骨碎片(A1-A3),左第一指骨(B1-B3)。C1-H2。Hippidion sp。左侧第一指骨(C1-C3: PIMUZ A/V 4205),左侧乳牙P2 (D1-D2: PIMUZ A/V 4239),左侧上颌磨牙碎片(E1-E2: MHNG GEPI V-3779),右下p3或p4 (F1-F2: PIMUZ A/V 4220),左下p3或p4和(G1-G2: PIMUZ A/V 4197),左侧M2 (H1-H2: PIMUZ A/V 4198)。视图:唇(D2, F2, G2),侧(C3),舌(E2, H2),掌/足底(B2, C2),右侧(A1),咬合(A3, D1, E1, H1, G1),近端(B1, C1)和腹侧(A2)。犬齿(c)、下锥体(hypc)、门齿(i)、中柱(msty)、后锥体(mtcd)、介柱(mty)、柱旁(prsty)、上颌骨(px)、原锥体(prtc)、上磨牙(M)和上前磨牙(P)
一个破碎的头骨,与中央跗骨相连的右侧距骨,以及来自同一个体的左侧第一指骨(PIMUZ A/V 4100)。标本的地理数据见附加文件1。
PIMUZ A/V 4100是一个仅保存在其腹侧的不完整头骨(图8A1)。前颌有左I3,两颌均有犬齿,有完整的左、右牙系列P1-P4和M1-M3。除P1外,前磨牙和磨牙强健,呈方形;前磨牙大于臼齿(图8A2-A3)。前磨牙的咬合长度大约在45到33毫米之间,磨牙的咬合长度在29到33毫米之间。右侧M1-M3系列长度为92 mm,左侧为94 mm。右侧系列P1-M3的总长度为216mm,左侧系列为217mm。前磨牙和磨牙的磨损程度不高,可以清楚地观察到牙齿的形态,其特征是卵形至圆形的原锥体,角状/近三角形的下锥体,副柱比中柱更明显。pre/ postfossets仅在M1中可见。其他可能属于同一个体的颅/颅后部分包括两个保存状态较差的颅骨碎片,一个处于碎片状态的右距骨,与中央跗骨相连,第三个手指的第一左指骨长度为85毫米(图8B1-B2)。
根据Alberdi等人(2005,2006)的研究,Hippidion的牙列具有椭圆形到圆形的原锥体的特征,这一特征在PIMUZ A/V 4100中也存在。Hippidion被认为是南美洲的特有属,其生物年代从上新世到晚更新世(Prado & Alberdi, 1996)。已经确认的有三个物种,(1)希皮迪翁(Hippidion principale),是最健壮的物种,在阿根廷的潘帕草原最为人所知,秘鲁和厄瓜多尔也可能有报道;(2) Hippidion devillei,这是一种中等大小的物种,化石记录遍布秘鲁安第斯山脉中部和阿根廷Pampa(尽管它可能远至委内瑞拉北部,见Labarca et al., 2021);(3)最小的物种——saldiasi Hippidion,仅存在于巴塔哥尼亚地区和智利北部一些地区的晚更新世沉积物中(见Alberdi & Prado, 1993;Labarca et al., 2021;Prado & Alberdi, 2017)。saldiasi Hippidion saldiasi是Roth于1899年命名的一种(原名onohippidium saldiasi Roth, 1899)。河马物种之间的区分结合了对头骨和跖骨特征的分析,重点是尾骨的大小和比例(Alberdi & Prado, 1993;Alberdi et al., 1995;Prado & Alberdi, 2017)。PIMUZ A/V 4100具有与阿根廷、巴西和玻利维亚Tarija的H. principale相似的牙齿特征(例如,Alberdi et al., 2001b, 2003,2006;MacFadden, 1997;Prado et al., 2012, 2013)。PIMUZ A/ v4100中M1-M3系列的尺寸和P1-M3系列的总长度符合H. principale的长度变异范围。将PIMUZ A/V 4100分配给最大的河马物种H. principale似乎是可能的。
Hippidion sp。
(图8 c1-h2)。
左上颌骨有四个臼齿和两个独立的右侧臼齿(MHNG GEPI V-3779),两个右侧M3 (PIMUZ A/V 4115, 4238),左侧dP2 (PIMUZ A/V 4239),左侧M2 (PIMUZ A/V 4198)和左侧M3 (PIMUZ A/V 4222),右侧p3或p4 (PIMUZ A/V 4220),左侧p3或p4和(PIMUZ A/V 4197),和p2或p3 (PIMUZ A/V 4219),右侧M3 (PIMUZ A/V 4240)和左侧第一指骨(PIMUZ A/V 4205)。标本的地理数据见附加文件1。
左侧上颌骨MHNG GEPI V-3779(图8E1-E2)除了两个分离的右侧P3或P4以及M1或M3外,还保留了四个臼齿(P3 - m2),可能来自同一个体。上颌磨牙的咬合长度大约在前磨牙32 - 33毫米之间,磨牙30 - 31毫米之间。前磨牙显示凹窝和一个突出的亚椭圆形原锥体(图8E1)。其他分离的上前磨牙和磨牙(PIMUZ A/V 4115, 4198, 4222, 4238-39;(见图8D1-D2, H1-H2)坚固,原锥呈椭圆形/椭圆形至圆形。这些臼齿形的咬合长度大约在27到45毫米之间。分离的下前磨牙/臼齿(PIMUZ A/V 4197, 4219, 4220, 4240;图8F1-G2)的特征是圆而小的元突和亚突,圆的原突和下突,这是河马的典型牙齿模式(Alberdi & Frassinetti, 2000;Alberdi & Prado, 2004;Alberdi等人,2001,2003,2005,2006,2007;Cerde?o等人,2008;MacFadden, 1997;Prado et al., 2012, 2013;Scherer & Da Rosa, 2003)。下离体臼齿的咬合长度大约在30到37毫米之间。第三指第一指骨(PIMUZ A/V 4205,图8C1-C3)保存状态不佳,最大长度为80mm,宽度为64mm。
MHNG GEPI V-3779的磨牙形具有凹窝和一个突出的亚椭圆形原锥体,不同于此处分配给Hippidion cf. H. principale的PIMUZ a /V 4100的磨牙形中的椭圆形到圆形原锥体(见图8A3)。我们不能排除来自潘潘地区的molariforms MHNG GEPI V-3779可能属于H. principale或H. devillei。saldiasi河马体型较小,迄今为止仅限于巴塔哥尼亚地区的晚更新世和智利的一些地方(Labarca et al., 2021;Prado & Alberdi, 2017)。我们的比较不允许提供更精确的确定这些标本,因为它们的碎片性质和先进的牙齿磨损(例如,PIMUZ a /V 4115, 4198, 4219- 20,4222, 4238-40)。参考左第一指骨PIMUZ A/V 4205,其宽阔的形状表明属于Hippidion sp.(见Alberdi et al., 2001a)。
林奈(1758年
隆德新马,1840年
马属新马属
(图9 a1-e2)。
图9
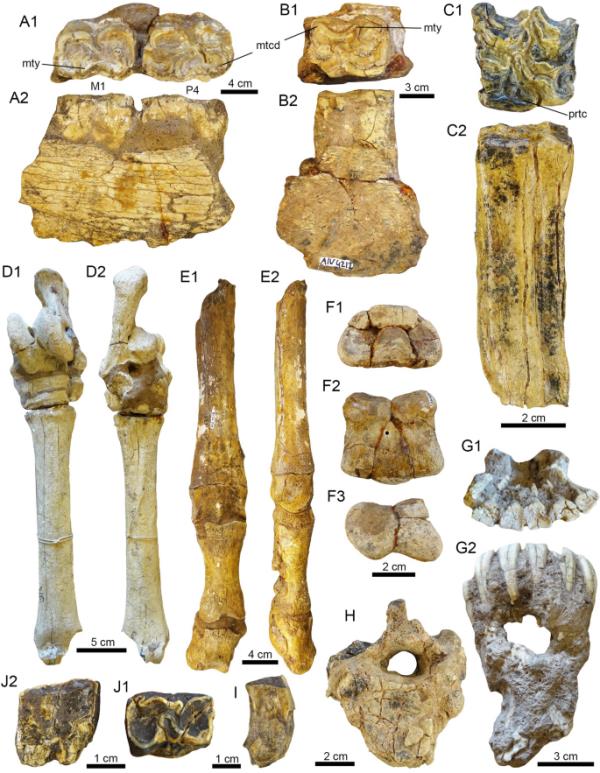
潘潘地区的马科动物。A1-E2。马属新马属a1 a2。半吞食的残片?p4-m1 (PIMUZ A/V 4183)。b1 b2。带有m1 (PIMUZ A/V 4212)的半食用片段。c1。左M1 (PIMUZ A/V 4236)。D1-D2。左跖骨(MTIII)具关节跗骨(MHNG GEPI V-3778)。E1-E2。右掌骨(MCIII)与第一和第二指骨(PIMUZ A/V 4248)。F1-J2。马科不定。F1-F3。第三指的第二指骨(PIMUZ A/V 4242)。G1-G2。下节骨与门牙(MHNG GEPI V-3780)。H.胸椎(PIMUZ A/V 4132)。1 .门牙(PIMUZ A/V 4241)。J1-J2。不确定下磨牙(PIMUZ A/V 4244)。视图:背侧(D1, E1, G1),右侧(E2),唇侧(B2),侧侧(F3),舌侧(A2, C2, J2),咬合(A1, B1, C1),掌/足底(D2, F2),后侧(H),近端(F1),腹侧(G2), Indet。(1). metaconid (mtcd), metastylid (mty), protocone (prtc)
左M1 (PIMUZ A/V 4236),左p4-m1 (PIMUZ A/V 4183)和左M1 (PIMUZ A/V 4212),不完整的右掌骨(MCIII)伴第一和第二指骨(PIMUZ A/V 4248),不完整的左跖骨(MTIII)伴关节跗骨(MHNG GEPI V-3778)。标本的地理数据见附加文件1。
PIMUZ A/V 4236是一个相对完整的左M1(图9C1-C2),缺少中部骨质,咬合长度约为30 mm。咬合面保存完好,可以观察到pli caballin和一个定义明确的三角形原锥体,其远端比中端长。左下p4-m1 (PIMUZ A/V 4183,图9A1-A2)的咬合长度分别为26 mm和24 mm。左m1 (PIMUZ A/V 4212,图9B1-B2)咬合长度为23 mm。标本的咬合表面保存完好,显示出双结,其特征是圆形的元突和相对有角的亚突,延长的外屈体,以及原锥体和下锥体,唇缘略直(见Alberdi等人,2002;Prado & Alberdi, 2017)。
标本PIMUZ A/V 4248是一个不完整的右MCIII,缺少其近端部分,并与第一和第二指骨相连(图9E1-E2)。由于这些骨头是通过物理连接保存下来的,我们无法进行精确的测量,但它的最大长度约为83毫米。明显的骨骺线表明骨干和骨骺之间的融合不完全,这表明PIMUZ A/V 4248可能属于亚成年个体。MHNG GEPI V-3778以左侧MTIII为代表,缺少远端骨骺左侧部分,因此估计总长度为240 mm;所有跗骨仍能连接,包括距骨和跟骨(图9D1-D2)。
如上所述,PIMUZ A/V 4183、4212和4236中存在的牙齿咬合模式和特征(例如,原锥体、元锥体、pli caballin)可能与马属动物的关系比与Hippidion的关系更大(参见Prado & Alberdi, 2017,以及其中的参考文献)。第一个与右MCIII相连的方阵(PIMUZ A/V 4248)比希皮皮顿的方阵更细长;其最大长度约为83毫米,与马属相似(见Prado等人,2012)。同样,左侧MTIII (MHNG GEPI V-3778)相当细长的形态是马属的特征,其最大长度(240毫米)与新马属的估计值相似(见Prado et al., 2012,表1)。
在过去的几十年里,南美马属化石的分类有效性一直存在争议(见Alberdi & Prado, 2004;Prado & Alberdi, 1994,2017)。Hoffstetter(1950)将来自南美洲的所有不同种类的马都纳入了Amerhippus亚属,仅基于下门牙表面缺乏凹痕和随之而来的表面牙釉质损失;然而,奥兰多等人(2008)和普拉多和阿尔伯迪(2017)对Amerhippus的有效性和使用提出了质疑。传统上,在南美洲的化石记录中发现了五个马属物种(例如,Prado & Alberdi, 1994),但Prado & Alberdi(2017)基于对颅骨和颅骨后元素的形态学和形态计量学特征的广泛修订,认为只有三个物种是有效的(1)E. andium (?(2) E. insulatus(中晚更新世)和(3)E. negeus(晚更新世)。最近,Machado & Avilla(2019)对三种马属物种的诊断和分类有效性提出了质疑,这表明南美洲可能只有一种本土马属物种,即E. neogeus。我们不排除本文所描述的来自潘帕斯地区的标本由于它们的形态可能属于E. neogeus。
马科indet。
(图9 f1-j2)。
下节骨,有六个门牙(MHNG GEPI V-3780),右m1或m2 (PIMUZ A/V 4203),不确定的下磨牙(PIMUZ A/V 4244),两个分离的破碎磨牙和一个分离的门牙(PIMUZ A/V 4241),胸椎(PIMUZ A/V 4132),第三指的第二个指骨(PIMUZ A/V 4242)。标本的地理数据见附加文件1。
MHNG GEPI V-3780(图9G1-G2)显示下合骨有6个门牙,牙合面磨损严重,可能是老年人。该标本连同其他分离的磨牙形(PIMUZ A/V 4203、4241、4244,图9I-J2)、胸椎(PIMUZ A/V 4132,图9H)、第三指的第二指骨(PIMUZ A/V 4242,图F1-F3)均呈碎片状或保存状态较差。
上述标本的残缺和/或保存状况较差,使其难以确定诊断特征以进行更精确的鉴定。
下载原文档:https://link.springer.com/content/pdf/10.1186/s13358-023-00273-7.pdf








